Mycoviruses, which infect fungi, are ubiquitous across all major fungi taxa, typically manifest as latent infection and seldom cause apparent symptoms in their hosts [1–3]. Advances in high-throughput sequencing and metaviromic have greatly expanded our understanding of mycoviruses diversity, revealing that a single fungal strain can be co-infected by multiple viruses. Interestingly, some mycoviruses enhance the virulence of their hosts, others attenuate pathogenic traits, thereby modulating the host biology. These alterations may include irregular colony morphology, reduced growth rate, abnormal pigmentation, impaired sporulation, and changes in sexual reproduction [4, 5]. The most concerning effect is mycovirus induced hypovirulence, which infection attenuates the pathogenicity of plant pathogenic fungi. This phenomenon presents mycoviruses as promising biocontrol agents for plant diseases [6, 7].
Mycoviruses with linear double-stranded RNA (dsRNA) genomes are currently classified into eight families: Amalgaviridae, Chrysoviridae, Megabirnaviridae, Partitiviridae, Quadriviridae, Reoviridae, Totiviridae, and Polymycoviridae (ICTV, https://talk.ictvonline.org/ictv-reports/ictv_online_report/). The family Paritiviridae currently comprises of five established genera: Alphapartitivirus, Betapartitivirus, Cryspovirus, Deltapartitivirus, and Gammapartitivirus, along with two proposed genera, Epsilonpartitivirus and Zetapartitivirus [8, 9]. With the except of Cryspovirus and Deltapartitivirus, members of the other genera have been reported to infect fungus. Partitiviruses are small, isometric, non-enveloped viruses with bisegmented dsRNA genomes ranging from 3.0 to 4.8 kbp in size [8]. The two essential genome segments, dsRNA1 and dsRNA2, typically encode the RNA-dependent RNA polymerase (RdRp) and coat protein (CP), respectively [10]. Occasionally, partitiviruses harbor defective interfering RNA (DI-RNA) that can modulate viral infection symptom in its hosts. For instance, Rosellinia necatrix partitivirus 2 (RnPV2) carries a DI-RNA that alters viral induction in Cryphonectria parasitica [11]. Similarly, an additional dsRNA segment has been identified in also found in certain isolates of Aspergillus flavus partitivirus 2 [12, 13]. The fungus infected by partitiviruses are generally asymptomatic, However, some members, such as Sclerotinia sclerotiorum partitivirus 1 (SsPV1/WF-1), Botrytis cinerea partitivirus 2 (BcPV2), and Colletotrichum liriopes partitivirus 1 (ClPV1), have been reported to reduce host virulence, mycelial growth, or conidial production [14–16]. beyond fungi, partitiviruses like the Osugoroshi viruses (OGVs) are associated with male-specific mortality after hatching in oriental tea tortrix, Homona magnanima [17]. Additionally, partitiviruses can exhibit broad host adaptability. A notable example is Penicillium aurantiogriseum partiti-like virus 1 (PaOLV1), which can stably replicate in a new fungal host, Cryphonectria parasitica, and confers enhanced resistance to salinity stress [18].
Fusarium oxysporum is an important plant pathogenic fungus that causes Fusarium wilt and Fusarium root rot in many crops worldwide [19–21]. Compared to other pathogenic fungi, relatively few mycoviruses have been reported in F. oxysporum. The identified dsRNA viruses belong to the family Chrysoviridae, Alternaviridae, Polymycoviridae, and Partitiviridae, as well as the proposed genus Unirnavirus [22–26]. Reported positive-sense single-stranded RNA [(+)ssRNA] viruses fall within the family Hypoviridae, Botourmiaviridae, and Mitoviridae [27–30]. One negative-sense single-standed RNA [(-)ssRNA] virus from family Mymonaviridaehas been described [31]. Co-infection of a single fungal strain by multiple viruses is common in species such as Sclerotinia sclerotiorum, Magnaporthe oryzae, Rosellinia necatrix, and Macrophomina phaseolina [32, 33]. However, the co-infection of F. oxysporum by two distinct viruses has not been previously reported.
In our previous studies, we found two dsRNA viruses, designated Fusarium oxysporum virus 1 (FoV1) and Fusarium oxysporum partitivirus 1 isolate 3S-18 (FoPV1/3S18) co-infect a single strain 3S-18 of F. oxysporum. We only reported the genomic characteristics and phylogenetic relationships of FoV1, but its effects on the host have not yet been investigated [25]. In this study, we analyzed the molecular characteristics and phylogenetic relationships of the other virus FoPV1/3S18. Furthermore, we demonstrated that FoV1 is the core virus responsible for hypovirulence in the host, and further evaluated the potential of FoV1 as biocontrol agents against Fusarium root rot.
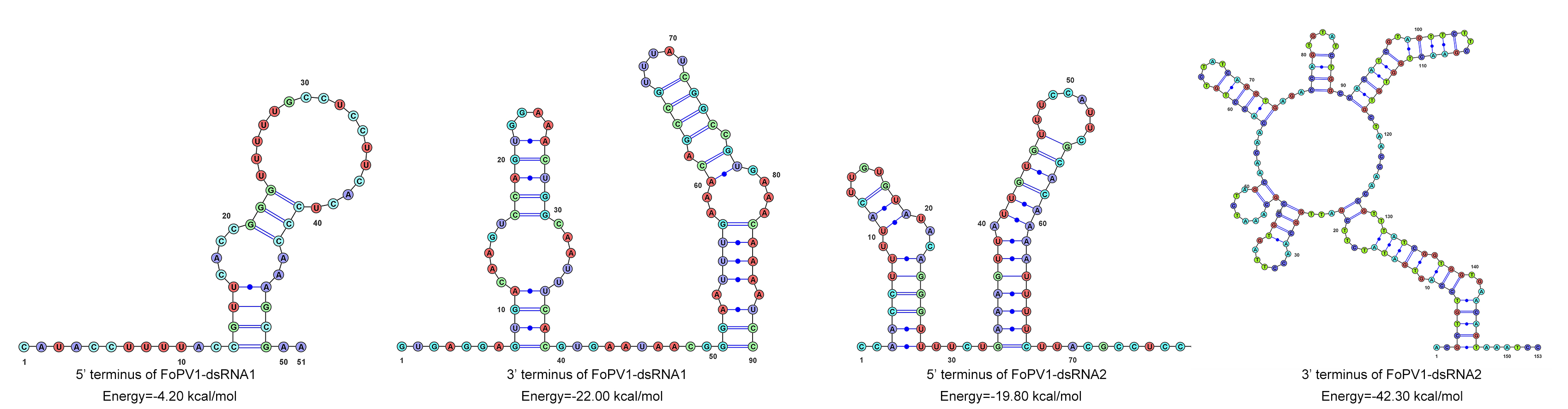{kind=link}