The 32 plant rhabdovirus sequences reported here were independently identified by 18 collaborating institutes/universities, each using different HTS approaches. Most species were identified by only one institute, whereas a few were identified by multiple institutes. Our study illustrates not only the diversity of plant rhabdoviruses but also the practical benefits of pre-publication data sharing for accelerating virus discovery, characterization, and contextualization. This collaborative approach reduced duplication of efforts, offered early insights into host range, geographical distribution and potential symptom associations, all of which support taxonomy and pest risk assessments [11, 13, 14, 28]. Such coordinated efforts also increase transparency and encourage data reuse, thereby advancing the field of plant virology.
Since most samples with virus-like symptoms were coinfected with other viruses, it was not possible to determine whether the identified rhabdoviruses are associated with symptoms. As Fox [12] emphasizes, establishing a causal relationship in plant virology is often challenging, particularly in mixed infections. Moreover, in several cases, it remains uncertain whether the virus-like symptoms were induced by viruses at all or by other factors. Further biological characterization studies, ideally using singly infected plants in controlled conditions, will therefore be required to determine potential etiological relationships.
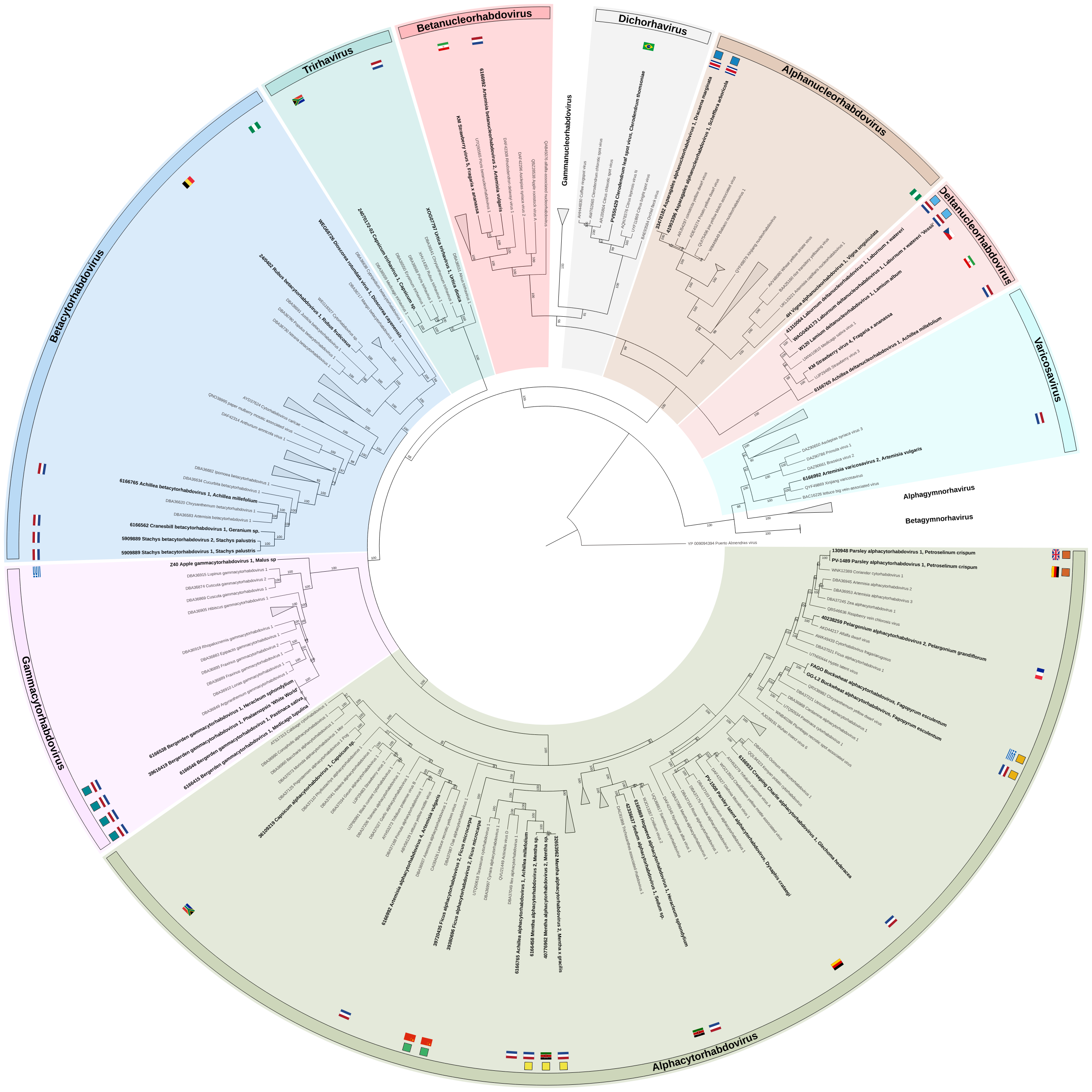
Nevertheless, two examples suggest potential virus-disease associations involving singly-infected samples. Laburnum deltanucleorhabdovirus 1 was detected in two Laburnum × watereri samples. The viral sequence was found both in a symptomatic herbarium specimen collected in 1967 where it occurred as a single infection, and in a living Laburnum × watereri tree co-infected with Arabis mosaic virus (Nepovirus arabis). Both plants exhibited similar virus symptoms of vein-yellowing and vein-banding with the living tree also showing mosaic patterns (Fig. 3a,b). Historical records by Masters [23] in 1877, van Katwijk [30] in 1953, and transmission electron microscopy observations of rhabdovirus-like particles by Cooper [8] support a long-observed potential link between vein-banding and mosaic symptoms in Laburnum and virus infection. This case also demonstrates the value of integrating historical herbarium material with modern molecular techniques. Similarly, Clerodendrum leaf spot virus was detected in singly infected Clerodendrum thomsoniae plants (data not shown), exhibiting chlorotic leaf spots (Fig. 3c), indicating potential pathogenicity of this virus. For both examples additional studies are needed to establish potential etiological relationships, ideally following the integrated approaches of Fontdevila Pareta et al. and Fox et al. [11, 12], including but not limited to screening of both asymptomatic and symptomatic plants in ecosystems and inoculation in controlled conditions.
In addition to 20 symptomatic plant samples, our study included 13 asymptomatic samples in which putative novel rhabdoviruses were identified. Many of these asymptomatic samples originated from virus reservoir surveys in wild plants, suggesting that numerous rhabdoviruses may not induce obvious symptoms in their hosts [5]. This is consistent with reports from other virus families, where asymptomatic infections are also frequently observed [25–27]. Together, these findings illustrate the high viral diversity that can infect apparently healthy plants within and outside agricultural ecosystems and supports the view that large-scale virus reservoir studies are important for biosecurity as they provide insights into the host range of viruses and allow better identification and allocation of the species potentially posing a phytosanitary risk [13].
Same rhabdovirus repeatedly detected in the same host species
Pre-publication data sharing enabled the early detection and cross-validation of potential virus–host associations and revealed that certain putative virus species are found across different countries. For example, parsley alphacytorhabdovirus 1 was independently detected in Petroselinum crispum (parsley) samples from the United Kingdom and Germany. Similarly, buckwheat alphacytorhabdovirus was identified in Fagopyrum esculentum (buckwheat) growing in habitat-enhanced field margins in Greece and the Netherlands. In addition, strawberry virus 4 and strawberry virus 5 were detected in the USA and Iran, suggesting a broad geographic presence. Ficus alphacytorhabdovirus 2 was detected in two Ficus microcarpa plants imported separately from China, cross-validating its host and distribution. Furthermore, Mentha alphacytorhabdovirus 2 was detected in three samples, namely from two cultivated and one wild Mentha species from both the Netherlands and Kenya. These examples highlight the practical value of data sharing, which allowed the independent identification of similar virus genomes in the same host across multiple countries, suggesting these viruses have been circulating for a long time or spreading between countries, for example through international trade.
Multiple rhabdoviruses infecting the same host species
In some plant samples, multiple distinct rhabdoviruses co-occurred. Stachys betacytorhabdovirus 1 and Stachys betacytorhabdovirus 2 were found in a single Stachys palustris plant (sample 5909889), while four distinct alphacytorhabdoviruses were identified in bulked Artemisia vulgaris (sample 6166992): Artemisia alphacytorhabdovirus 1–4. Similarly, in bulked sample Achillea millefolium (sample 6166765), both Achillea alphacytorhabdovirus 1 and Achillea betacytorhabdovirus 1 were identified, as well as Achillea deltanucleorhabdovirus 1, although only its L gene was assembled. These observations highlight the substantial rhabdovirus diversity that can exist within a single host.
Same rhabdovirus in different host species
Two rhabdoviruses were identified in more than one host species. Asparagales alphanucleorhabdovirus 1 was identified in a Heptapleurum arboricola and a Dracaena marginata plant, both imported from Costa Rica. Although both plant species belong to the same order (Asparagales), they are members of different families. Similarly, Bergerden gammacytorhabdovirus was identified in three asymptomatic wild species from a single location (Bergerden) and in a symptomatic, cultivated Phalaenopsis orchid. These findings suggest that both viruses may be transmitted by a polyphagous vector and that further screening may reveal additional host plant species, as observed for Physostegia chlorotic mottle virus (PhCMoV; Alphanucleorhabdovirus physostegiae) [28, 29].
Hidden diversity of plant rhabdoviruses
In the past decade, many plant rhabdoviruses have been identified through diagnostic testing, virus reservoir studies and mining of plant transcriptome database studies [3, 5]. However, as with other virus families, many findings are not being formally reported due to time constraints and because priority is often given to viruses or virus groups with clear phytosanitary impact [11, 13]. Our data-sharing-based approach led to the collective identification and publication of 32 putative novel species, underscoring the hidden diversity of this virus group.
Bejerman, et al. [3] reported 27 novel rhabdoviruses through SRA mining, roughly half of which were (putative) cytorhabdoviruses. Similarly, 63% (20 out of 32) of the putative novel rhabdoviruses presented in our study, not identified from the SRA but from actual plant samples, were also cytorhabdoviruses (including alpha-, beta- and gammacytorhabdoviruses). This suggests a rich, but underexplored diversity within this cytorhabdoviruses. However, it is important to note that a large diversity may also exist in other rhabdovirus groups but that this diversity is yet uncovered for example due to under sampling. Gymnosperm-infecting alpha- and betagymnorhavirus, for instance, are likely underrepresented, as gymnosperms tend to be sampled less than herbaceous plant species [3].
This study accounts for nearly 12.6% of the currently known plant rhabdoviruses species and makes a substantial contribution to the family diversity.
Virus discovery versus biological characterization in the HTS-era
With HTS now available to many labs, the challenge has shifted from virus discovery to the biological characterisation of these putative new viruses. This is due to the associated time-consuming efforts of biological characterisation, with priority typically given to findings with clear crop/plant health or phytosanitary impacts, leaving other findings unreported and dormant on servers [13, 17]. In addition, large amounts of neglected or unused data await secondary analysis and repurposing. Bejerman, et al. [2] predicted that the increasing use of HTS would result in the identification of many more novel viruses with negative-sense and ambisense RNA, including members of the family Rhabdoviridae, which is underlined by the 32 novel viruses described here. Although only limited biological, epidemiological and contextual data were available for most of the putative novel viruses in our study, we believe that reporting our findings will encourage other researchers to examine their dormant sequences and datasets. Additionally we hope it will inspire virus reservoir studies, including on asymptomatic plants, and prompt researchers to make their findings publicly available. This would increase our knowledge on host range, distribution, vectors, symptomatology, phytosanitary risks and general understanding of virus epidemiology.
Beyond motivating individual research efforts, our study shows the value of pre-publication data sharing as an important part of plant-health preparedness. Such sharing supports regional and global cooperation and rapid response and is similar to frameworks like ‘disaster plant pathology’ [10], the global crop disease surveillance system proposed by Carvajal-Yepes et al. [7], and parallel initiatives in animal and human virology, such as the Global Virus Network (https://gvn.org/).
In this sense, our work goes beyond filling taxonomic gaps and may contribute to informing the development of more coordinated and responsive approaches for plant-virus monitoring in the future.
{kind=link}