Diethyldithiocarbamate (DDC) is a well-characterized copper-binding compound that has been historically used for heavy metal chelation in both industrial and clinical settings, with increasing interest in its anticancer potential when complexed with copper[36, 37]. Given recent evidence that copper complexes of DDC can induce immunogenic cell death (ICD)[38], we first evaluated whether DDC promotes ATP release, a hallmark of ICD, when paired with transition metals (Fig. 1A–C). DDC alone had a minimal effect, but in combination with various copper salts, it triggered a 3- to 9-fold increase in extracellular ATP. Other transition metals such as cobalt, iron, or zinc did not show a comparable effect. The terpyridine analog TTBT, tested in parallel, strongly induced ATP as a single agent (~ 8-fold), with the copper combination further enhancing this to 30- to 40-fold. In contrast to copper, most other metals suppressed TTBT's ATP-releasing activity of TTBT, with the exception of cobalt, which had no effect. Transition metals alone had a limited effect on ATP release (Fig. 1C). Dose-response analysis confirmed that ATP release by CuDDC was concentration-dependent and detectable at concentrations as low as 3.1 µM in B16-F10 cells (Fig. 1D–E).
To better understand whether this ATP induction was primarily due to copper transport or the intrinsic biological effects of the ligands, a panel of copper-binding compounds from several drug classes was profiled (Fig. 1F). Cytotoxicity was assessed with and without CuSO₄ at 24 and 72 hours. Although the IC₅₀ values varied widely, copper potentiation, defined as a shift in IC₅₀, was only observed for DDC, 8-hydroxyquinoline (8HQ), and pyrithione, which are known ionophore-like compounds[39]. To assess whether this potentiation correlated with copper transport, cells were pulsed for 4 hours with each compound and CuSO₄ and the intracellular copper levels were measured (Fig. 1G). Ionophore-like compounds significantly increased intracellular copper levels (2- to 8-fold). The terpyridine analogs TTBT and CPT also increased the copper content by 3- to 5-fold, respectively. Other compounds, including several polyphenols and chemotherapies known to bind divalent metals, had minimal effects or slightly decreased copper accumulation.
These findings indicate that copper ionophore-like compounds significantly enhance intracellular copper accumulation and potentiate cytotoxicity in the presence of extracellular copper. While some compounds such as TTBT, CPT, and clioquinol increased copper levels without corresponding increases in cytotoxicity, others such as DDC showed time-dependent potentiation, with effects observed at 24 h but not at 72 h.
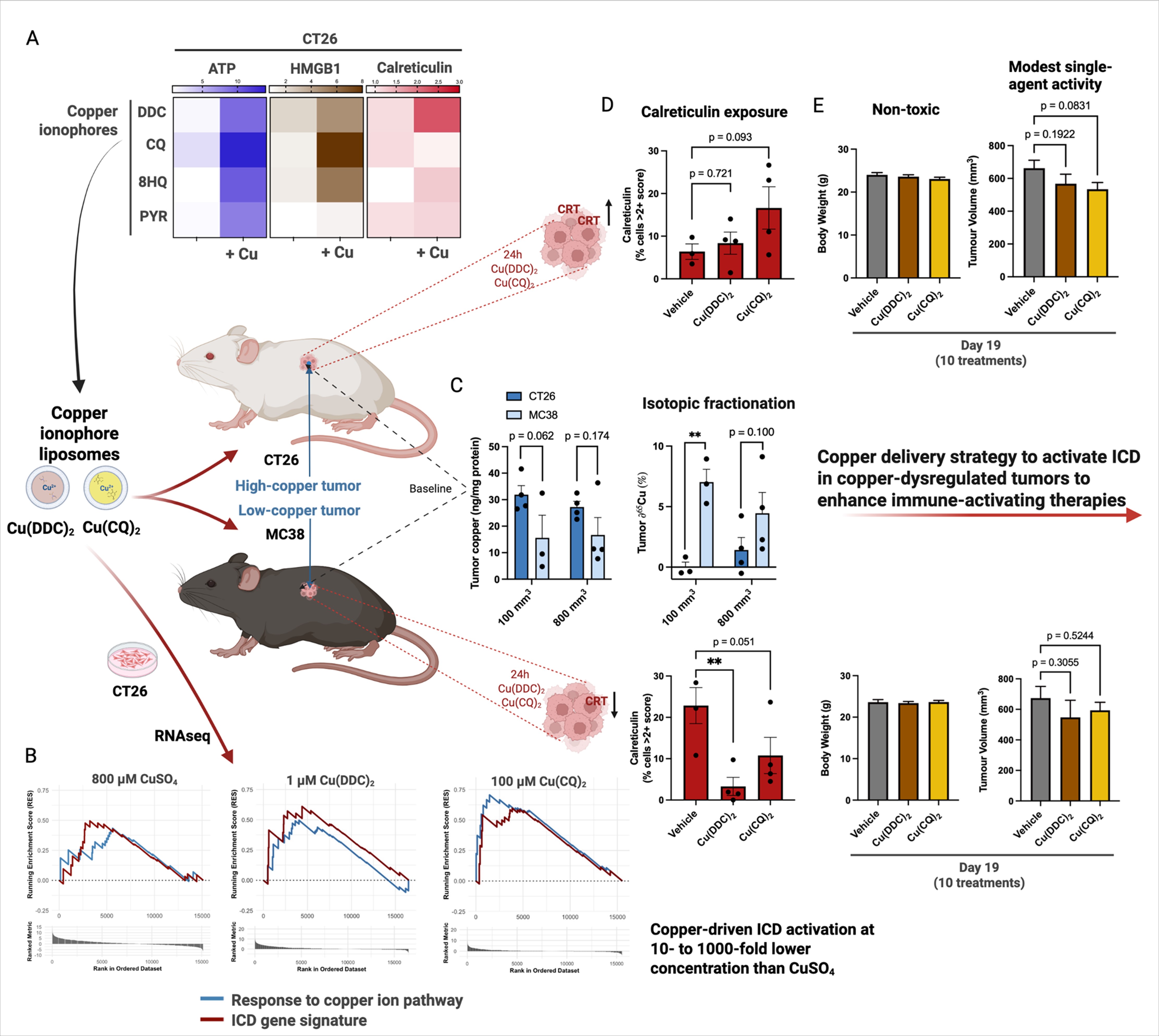
To evaluate whether copper-ligand combinations promote immunogenic signaling, three damage-associated molecular patterns (DAMPs)–ATP release, HMGB1 release, and calreticulin (CRT) exposure–were measured in CT26 cells following 24-hour treatment (Fig. 2A–C). The compounds were applied at doses optimized for ATP induction as single agents (Fig. S1), either alone, with CuSO₄, or CuSO₄ alone at the same concentration. The known ionophore group (brown bar) showed minimal activity across DAMPs when used alone, but the addition of copper markedly enhanced all three markers, as reflected in the fold-increase column. In contrast, compounds in the terpyridine class (pink bar) induced DAMP release as a single agent with little further enhancement upon copper addition. TTBT was the only terpyridine compound that increased all three DAMPs in the presence of copper. The other compound classes showed minimal effects regardless of the addition of copper.
To determine whether intracellular copper levels alone accounted for DAMP induction, internalized copper from Fig. 1G was compared to the DAMP responses following CuSO₄ treatment alone (CuSO₄ columns in Fig. 2A–C). There was a strong positive correlation between the intracellular copper content and the release of ATP, HMGB1, and CRT (Fig. 2D), suggesting that copper accumulation can directly stimulate ICD-associated pathways. The ionophore-like activity, defined as the fold increase in intracellular copper over CuSO₄ alone, was assessed to determine whether copper-induced enhancement of DAMP markers could be predicted. Ionophore-like activity was associated with HMGB1 and CRT release and showed a strong trend with ATP (Fig. 2E). However, when analyzing all treatments, there was no consistent correlation between total intracellular copper levels and DAMP release, likely because of copper-independent effects in some compound classes. Together, these results support a model in which copper ionophore-like activity drives ICD-like responses by increasing intracellular copper concentrations beyond the threshold achieved with soluble copper alone.
With intracellular copper delivery emerging as a potential strategy to activate ICD and enhance immune responses, copper uptake following addition of CuSO₄, Cu(DDC)₂, and Cu(CQ)₂ was assessed across a panel of human and murine cell lines (Fig. 3A–C). Baseline intracellular copper levels were measured, followed by a 4-hour treatment with each compound. Addition of Cu(DDC)₂ and Cu(CQ)₂ resulted in increases in intracellular copper when compared to cells incubated with 100 µM CuSO₄. The copper doses for the Cu(DDC)₂ and Cu(CQ)₂ studies were 20- and 4 fold less, respectively. To determine whether basal copper levels were predictive of treatment sensitivity, eight cancer cell lines were assessed, and cytotoxicity after 72-hour exposure to Cu(DDC)₂ and Cu(CQ)₂ was determined (Fig. 3E–F). A heatmap of cuproptosis-related gene expression revealed notable heterogeneity in core regulators such as FDX1, DLAT, and PDHA1 across cell lines (Fig. 3D), with MDA-MB-231 cells displaying both high expression of multiple cuproptosis-associated genes and the greatest sensitivity to Cu(CQ)₂, consistent with its elevated basal copper levels. When IC₅₀ values were plotted against baseline copper levels, no correlation was observed for Cu(DDC)₂ (Fig. 3H), whereas Cu(CQ)₂ cytotoxicity showed a significant association with basal copper content (Fig. 3I).
To examine the transcriptomic effects of copper treatments, gene ontology enrichment analysis was performed with CT26 cells treated with CuSO₄, Cu(DDC)₂, or Cu(CQ)₂ at IC₇₀ for 24 hours, a dose selected to capture death-related signaling while limiting confounding from growth inhibition. Gene set enrichment was performed using log₂ fold-change–ranked genes and mouse GO gene sets (Fig. 4). Enrichment maps revealed that CuSO₄ primarily induced pathways related to ion transport along with smaller clusters associated with cell motility and keratinization (Fig. 4A). Cu(DDC)₂ and Cu(CQ)₂ also showed enrichment in ion transport-related terms, likely reflecting their role in facilitating copper uptake; however, these agents had broader transcriptional responses that suggested changes in immune and cellular regulatory processes. Cu(DDC)₂ treatment led to enrichment of pathways related to membrane organization, vesicle trafficking, and hormone transport (Fig. 4C–D), potentially reflecting intracellular responses to altered metal homeostasis. Cu(CQ)₂, on the other hand, induced strong upregulation of immune-related pathways, including cytokine production, Toll-like receptor signaling, and regulation of adaptive immune responses (Fig. 4E–F). Some enriched categories also included terms associated with tolerance induction and suppression of lymphocyte activation. While the directionality of these responses remains uncertain, the pattern is consistent with the activation of both immunostimulatory and regulatory programs. These results indicate that, while the core transcriptomic effects of CuSO₄ and both ionophores involve metal ion processing, Cu(DDC)₂ and Cu(CQ)₂ engage distinct gene expression programs that reflect different cellular responses to copper delivery.
To determine whether transcriptional responses to copper accumulation corresponded with immunogenic stress signaling, the expression changes in Cu-responsive and ICD-related genes were profiled in CT26 cells. All three treatments activated genes associated with the Gene Ontology category "response to copper ion," with the most pronounced expression in CuSO₄ and Cu(DDC)₂, despite Cu(DDC)₂ being administered at an 800-fold lower molar dose compared to CuSO₄ (Fig. 5A). This suggests that intracellular delivery mechanisms may be more influential than the total amount of copper administered to drive the transcriptional response. Under the same conditions, the expression of a curated 31-gene immunogenic cell death (ICD) signature (Fig. 5B) was examined. Each treatment upregulated subsets of ICD genes but with differing breadths and intensities. Volcano plots showed that CuSO₄ induced widespread upregulation and downregulation of genes, which was consistent with a broad stress response (Fig. 5C). In contrast, Cu(DDC)₂ triggered more selective transcriptional activation, including strong upregulation of NLRP3 and TNF (Fig. 5D). Cu(CQ)₂ had fewer differentially expressed genes overall but included increased expression of P2RX7 and NLRP3 (Fig. 5E). These patterns suggest that CuSO₄ induces a broader transcriptional response, whereas ionophore-based treatments result in a more selective activation of immune-related gene pathways (Fig. 5E).
Gene set enrichment analysis (GSEA) plots visualized the overall enrichment of the copper response and ICD pathways (Fig. 5F–H). Although CuSO₄ induced the most differentially expressed genes, its enrichment for the copper ion response set was the weakest, likely because of widespread transcriptional changes that dilute pathway-specific signals. Cu(CQ)₂ yielded the strongest and most selective enrichment of both Cu-responsive and ICD-associated genes, despite lower overall transcriptional activation. Cu(DDC)₂ showed intermediate behavior, with strong activation of ICD components and robust copper-responsive transcription. These findings support a model in which ionophore-based copper delivery can modulate gene expression in a more targeted manner than extracellular copper alone, potentially shaping.
As shown in Fig. 3, CT26 cells in vitro had significantly higher intracellular copper levels than MC38 cells. Since this may affect the therapeutic response to copper delivery, copper and immune-related features were characterized in syngeneic murine tumor models following s.c. injection of the two cell lines (Fig. 6A). Copper levels in CT26 cells were approximately twice the level in MC38 cells, but this difference was less pronounced in larger tumors (Fig. 6B). The common isotopes of copper are ⁶³Cu and ⁶⁵Cu, and copper isotopic fractionation is thought to reflect cellular metabolism and transporter activity[41–43]. While altered ⁶⁵Cu/⁶³Cu ratios have been observed in cancer patients, recent data suggest that measurable isotopic shifts in serum may originate in the liver rather than directly in tumor tissue[44]. In this study, MC38 tumors showed higher ⁶⁵Cu enrichment than CT26 tumors, particularly smaller tumors (Fig. 6C). Plasma levels of total and isotopic copper were not significantly different when plasma was obtained from tumor-bearing mice (Fig. 6F–G).
Large MC38 tumors have higher TNFα and IL-12 cytokine levels, which may reflect immune activation. In addition, elevated IL-10 levels were observed, which may be associated with immunosuppression. Interestingly, plasma cytokine levels showed an opposite trend, with higher TNFα and IL-12 levels in CT26 tumors and lower levels in MC38 tumors (Fig. 6D, H). CT26 tumors also showed higher CD8 + infiltration than MC38 tumors, suggesting greater immune priming in this model. Similar levels of other lymphocyte populations and macrophages were observed in these two tumor models (Fig. 6E).
To determine the in vivo susceptibility of copper ionophore-like compounds and their ability to activate ICD in these models, CT26- and MC38-bearing mice were treated with one dose of Cu(DDC)₂ or Cu(CQ)₂. CRT exposure was assessed 24 h later using IHC (Fig. 6I). In the CT26 tumor model, Cu(CQ)₂, delivering 0.80 mg/kg elemental Cu, caused an increase in CRT levels (albeit not significant), whereas there was little change in this model when Cu(DDC)₂ was given (delivering 0.25 mg/kg Cu; Fig. 6J). MC38 tumors had higher baseline CRT levels, but treatment with both ionophore-like compounds led to decreased CRT expression, including a significant (p < 0.05) 7-fold reduction when the mice were injected with Cu(DDC)₂ and a 2-fold decrease (p = 0.051) when the mice were injected with Cu(CQ)₂ (Fig. 6K). These findings indicate important differences in copper accumulation and immune-related responses between the two tumor models, with implications for how Cu-based therapeutics might differentially modulate ICD in distinct tumor contexts.
The activity of monotherapies with Cu(DDC)₂ and Cu(CQ)₂ liposomes was assessed in CT26 and MC38 tumor models. Mice were treated three days after tumor cell inoculation, and a total of 10 intraperitoneal doses were given over two weeks (Fig. 7A). Both agents produced modest growth inhibition compared to the vehicle controls, with Cu(CQ)₂ showing a slightly greater effect in the CT26 model (Fig. 7B–C, E–F). Body weight was unaffected by this dose schedule (Fig. 7D, G), and no significant adverse events were observed. These data suggest that copper ionophore monotherapy is well tolerated but modestly suppresses tumor growth. These results suggest that these agents may be better suited for use in combination regimens aimed at enhancing immunotherapy through copper-dependent or other immunomodulatory mechanisms.
{kind=link}