Farmed abalone typically require 3–5 years to reach the market size of 80 mm. Therefore, growth performance is one of the most important factors in the abalone industry [33]. Over past years, probiotics have gained increasing popularity as they provide an alternative source of beneficial microorganisms for aquaculture, which typically relies on terrestrial microbes [34]. Our research investigates the impact of probiotics supplementation on abalone growth performance, muscle nutritional composition, gut microbiota structure, and lipid metabolism, aiming to lay a groundwork for the application of probiotics in abalone farming.
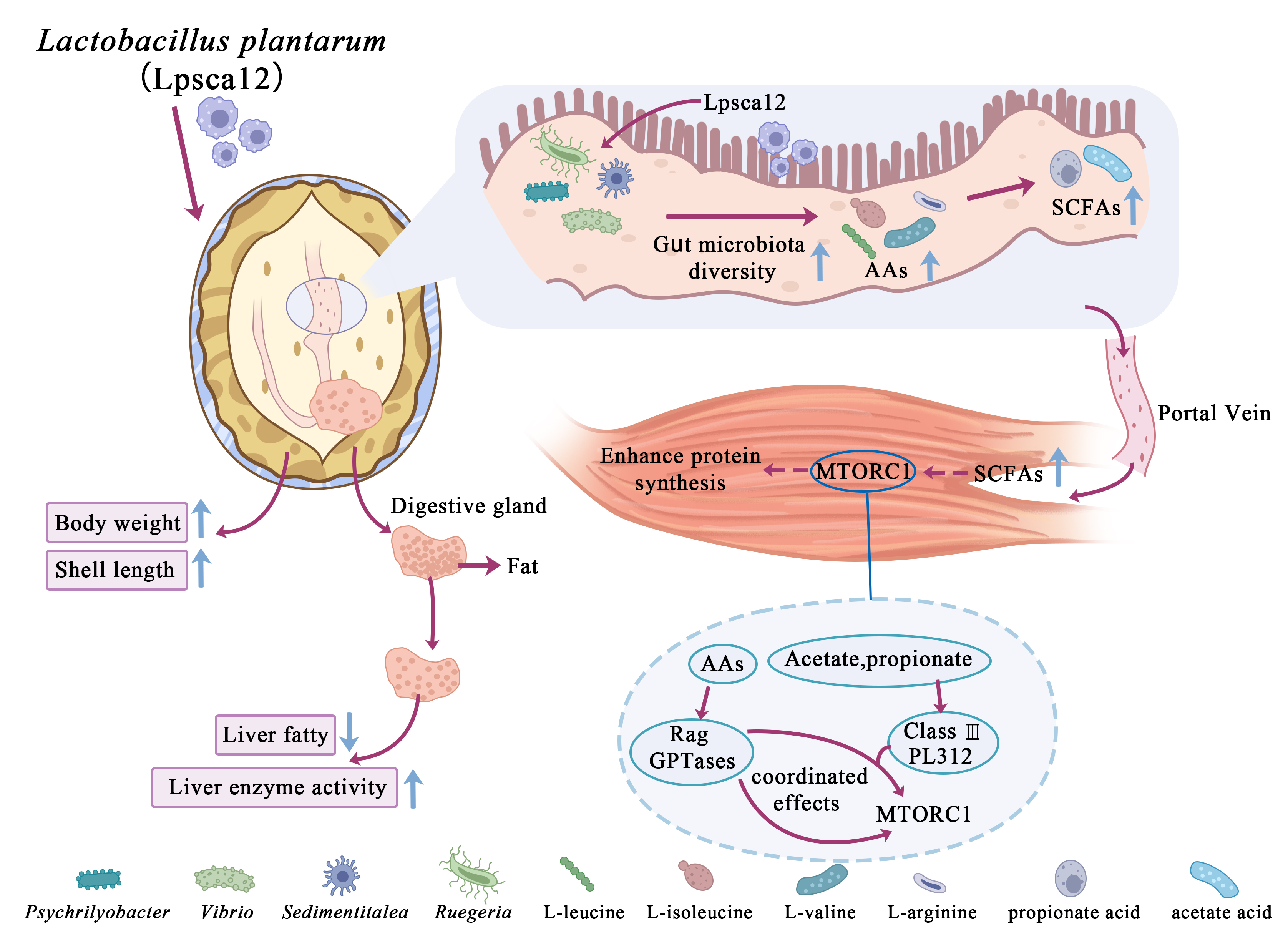
Our study found that LPsca12 treatment conferred beneficial effects on growth-related traits in abalone. In comparison with the Con group, body weight, shell length, and survival rate of the Pro group were significantly improved, which is in line with findings from studies on sea cucumbers and oysters [35, 36].
SCFAs, particularly acetate and propionate, play direct roles in muscle anabolism, these metabolites are primarily synthesized in the host intestine and subsequently transported via the portal circulation to peripheral tissues such as muscle, where they contribute to energy supply, inflammatory modulation, and muscle protein synthesis [37–39]. Acetate has been shown to activate muscle cell metabolism and promote myofiber formation through AMPK signaling, while Propionate induces mTORC1 pathway activation, which is a pivotal regulator of protein biosynthesis [40, 41]. However, studies on the effect of probiotics on SCFAs accumulation in abalone muscle remain limited. In our study, LPsca12 significantly elevated the content of acetate and propionate within abalone muscle tissue. Based on these findings, dietary LPsca12 may serve as a promising nutritional strategy to enhance muscle quality by elevating muscle-derived SCFAs levels and activating anabolic signaling pathways.
Fat accumulation in the liver is a prevalent metabolic disorder in fish aquaculture, significantly affecting their growth performance and immune capacity, and may also cause liver and pancreatic damage, as well as energy metabolism disorders [42, 43]. During the progression of lipid metabolism disorders, excessive circulating free fatty acids in adipose tissue are stored in the liver, leading to an imbalance in lipid homeostasis [44]. Elevated hepatic levels of TG and T-CHO are generally considered to be key features of lipid metabolism disorders and the formation of fatty liver [45]. Our study found that the levels of TG, T-CHO, and LDL-C in the liver of the Pro group were markedly reduced, indicating that hepatic lipid deposition was reduced, and lipid metabolism disorders were effectively improved. LPL, a rate-limiting enzyme in the metabolism of exogenous lipids, is an essential hydrolase that catalyzes the breakdown of triglycerides, releasing fatty acids for uptake and utilization by tissues [46]. HL regulates the liver's clearance andremodeling of lipoproteins and participates in tholesterol metabolism [47]. LPS functions in fat metabolism and transport by catalyzing the hydrolysis of lipids into glycerol and fatty acids [48]. The Pro group exhibited significantly higher hepatic LPL, HL, and LPS activity levels compared with the Con group, which helped to accelerate the clearance of exogenous lipids and optimize the overall lipid metabolism. Adding LPsca12 may improve the metabolic utilization efficiency of fatty acids by upregulating the expression and activity of liver lipases, thereby alleviating the lipid burden in the liver and promoting the restoration of lipid homeostasis.
As a major regulator, the gut microbiota influences intestinal immune function, nutrient uptake, and overall host health [24]. However, it has been reported that its composition and diversity are influenced by farming environments, including water salinity, pH, feed sources, and seasonality [49–51]. In our study, we applied metagenomics to investigate the gut bacterial communities in abalone simultaneously. The analysis revealed a statistically significant increase in alpha diversity in the Progroup relative to the Con group. Combined with β diversity analysis, we inferred that LPsca12 contributes to increased species richness as well as diversity of the abalone gut microbiota. Studies have shown that Firmicutes and Bacteroidetes constitute the predominant phyla present guts of healthy, contributing significantly to host health, immune function, and internal homeostasis [52, 53]. Our findings indicated that LPsca12 administration markedly elevated the abundance of Firmicutes and Bacteroidetes in the abalone intestinal microbiota. Additionally, some members of Psychrilyobacter and Vibrio were associated with healthy individuals [54, 55]. Psychrilyobacter has previously been reported as probiotics that can improve abalone survival and immunity [56]. Furthermore, functional annotation based on KEGG pathway analysis indicated that LPsca12 supplementation could modulate the host microbiota composition and facilitate the proliferation of beneficial microbes, potentially through suppression of pathogenic bacterial colonization within the gastrointestinal tract. These findings highlight that LPsca12 can positively regulate the abalone gut microbiota and promote host health.
Metabolomics offers an integrated view of metabolic profiles in biological specimens and enables detailed insights into host responses to diverse environmental stimuli and physiological conditions [57–60]. In our study, a total of 1,755 metabolites showed increased levels, while 341 exhibited decreased levels in the Pro relative to the Con group, and a notable enrichment of amino acids was observed in the Pro group. Beyond serving as essential substrates for protein synthesis, amino acids are critically involved in energy metabolism, neurotransmitter function, intestinal barrier maintenance, and the modulation of inflammatory responses [61, 62]. Furthermore, amino acid–derived metabolites (e.g., SCFAs, amines, phenolic compounds) function as key modulators of gut homeostasis and host–microbe interactions [63]. Previous evidence have indicated that intestinal microbiota can profoundly impact host metabolism and muscle protein synthesis through the regulation of amino acid metabolism [64]. However, the mechanisms by which probiotics regulate amino acid metabolism and their derivatives to influence muscle nutrition in abalone remain insufficiently understood. In our study, we identified a potential association between gut amino acid metabolism and SCFAs production; however, the underlying mechanisms and their effects on host physiology warrant further investigation.
Finally, we investigated the potential contribution of LPsca12 to muscle-related biosynthetic pathways. After probiotic supplementation, the intestinal microbiota of abalone exhibited significant enrichment, indicating an improved gut microbial environment. Based on this observation, we hypothesize that LPsca12 intervention modulated the microbial community, leading to detectable metabolic alterations. Notably, key nutrient metabolites, including branched-chain amino acids (L-leucine, L-isoleucine, and L-valine) and L-arginine, were significantly elevated in abalone tissues following probiotic treatment. Microbial catabolism of amino acids is known to generate ammonia, organic acids, and gaseous compounds, among which SCFAs are the most plentiful final metabolites [65]. This suggests that enhanced amino acid metabolism may effectively promote SCFAs biosynthesis, which is consistent with our findings. Among these amino acids, L-leucine and L-arginine are well-recognized activators of mTORC1 and may activate the mTORC1 complex via the Rag GTPase signaling pathway, thereby facilitating protein synthesis and myocyte growth, ultimately contributing to improved muscle nutritional status [66].In parallel, the elevated levels of SCFAs such as acetate and propionate in muscle tissue may activate the GPR41/43–AMPK–SIRT1 signaling axis, thereby enhancing mTORC1 responsiveness and synergistically regulating muscle energy metabolism and protein synthesis [67, 68]. Moreover, our study revealed that LPsca12 significantly improved hepatic lipid metabolic parameters, characterized by decreased TG, T-CHO, and LDL-C and increased LPS, LPL, and HL levels. These changes suggest enhanced lipid clearance capacity and reduce ectopic lipid deposition, which may contribute to a more favorable metabolic milieu for muscle protein anabolism.
Collectively, these findings suggest that LPsca12 may modulate muscle nutrition and metabolism in abalone through a “microbiota–AAs–SCFAs–mTORC1” signaling axis, possibly in concert with improved hepatic lipid metabolism. Although the precise molecular mechanisms remain to be elucidated, our study provides novel microecological insights for the regulation of muscle quality in aquatic animals.
{kind=link}