Bacillus mojavensis postbiotic selectively reduces the viability and migration of SW480 colon cancer cells without affecting PBMCs.
To determine the antiproliferative potential of B. mojavensis postbiotic, we first assessed its effect on SW480 colorectal cancer cells. Exposure to 25 or 50 µg/mL of the postbiotic for 24 h significantly reduced cell viability as measured by MTT assays (****p < 0.0001 vs. control), with both concentrations showing comparable effects. These findings were confirmed by crystal violet staining, which revealed a marked decrease in adherent cells and monolayer disruption, indicating loss of proliferative capacity and viability (Fig. 1A–B).
We next evaluated the impact on cell migration, a critical step in metastatic dissemination. Wound-healing assays showed that postbiotic treatment at both concentrations inhibited SW480 migration after 24 h compared to MRS control (Fig. 1C). This antimigratory activity may involve interference with cytoskeletal dynamics, extracellular matrix remodeling, or adhesion signaling pathways that sustain tumor invasiveness. Moreover, the effects on cell viability may be attributed to the metabolites secreted by B. mojavensis, regardless of the culture medium in which the postbiotic is generated. This is supported by the fact that when the postbiotic is produced in the DMEM F12 cell culture medium, it still decreases the viability of SW480 cells (Supplementary Fig. 1).
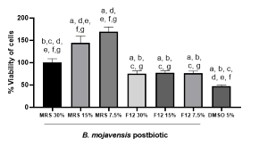
Given the therapeutic relevance, it was essential to determine whether these effects were selective for malignant cells or also detrimental to healthy immune cells. Therefore, PBMCs from healthy donors were exposed to postbiotic (30% v/v) or MRS control for 24 h. Interestingly, no cytotoxicity was observed in PBMCs. In fact, postbiotic-treated PBMCs displayed significantly higher viability compared to MRS (****p < 0.0001) (Fig. 1D). This result supports the selectivity of the postbiotic towards cancer cells, supporting its potential as a safe therapeutic adjuvant and ruling out general cytotoxicity. The inclusion of PBMCs was explicitly designed to evaluate biosafety, a critical requirement in developing biotherapeutic candidates.
Bacillus mojavensis postbiotic attenuates LPS-induced proliferative signaling in SW480 cells.
The tumor microenvironment of colorectal cancer is characterized by chronic inflammation, often exacerbated by bacterial products such as lipopolysaccharide (LPS), which engages Toll-like receptor 4 (TLR4) and activates NF-κB/MAPK cascades that promote tumor growth and survival. To model this inflammatory stimulus, SW480 cells were incubated with 10 µg/mL LPS for 24 h. As expected, LPS induced a strong proliferative response, increasing cell viability more than fourfold relative to control. Remarkably, co-incubation with B. mojavensis postbiotic (25 or 50 µg/mL) abrogated this effect, reducing proliferation by up to 375.68% compared to LPS alone (****p < 0.0001) (Fig. 1E). This finding indicates that the postbiotic may interfere with LPS-driven pro-inflammatory and proliferative cascades, although the precise molecular mediators remain to be identified.
Bacillus mojavensis postbiotic modulates oncogenic gene expression in SW480 cells.
Microarray analysis of SW480 cells exposed to 15% v/v postbiotic for 10 h revealed extensive transcriptional reprogramming. A total of 1,920 transcripts were interrogated, and the 20 genes with the highest positive (up-regulated) and negative (down-regulated) z-ratio values were represented in the heatmap (Fig. 2A). The heatmap illustrates the differential expression of these genes, with hierarchical clustering clearly separating the samples into two groups: control (MRS) and B. mojavensis postbiotic (BM). Red and blue colors indicate up- and down-regulated genes, respectively. The up-regulated genes included PLOD2, MMP14, RPS19, CD80, DCT, PRDX2, RAB10, ABCC4, SSTR2, and RCV1, while the down-regulated genes comprised PBOV1, SERPINB1, FLJ38507, OGN, RFXAP, MEST, SLC12A2, ENO1, HLA-DQA1, and RAD1.
In Fig. 2B, functional enrichment analysis of the 20 most representative DEGs is shown as a bar plot integrating multiple databases (GO, KEGG, Reactome, WikiPathways, miRTarBase, Transcription Factor, Human Protein Atlas, Corum, and Human Phenotype Ontology). Enriched categories are displayed on the X-axis and adjusted significance values (− log10 Padj) on the Y-axis. Only GO:CC (Cellular Component), GO:BP (Biological Process), and Transcription Factors (TF) showed significant enrichment. Among the most relevant terms were extracellular exosome, endocytic vesicles, transcription regulator complexes, and cell adhesion, highlighting the role of the postbiotic in modulating intercellular communication, transcriptional regulation, and vesicle organization. These findings suggest that treatment with B. mojavensis postbiotics alters both the dynamics of exosome secretion and gene regulation mediated by transcription factors that are critical in cancer, paving the way for future studies to identify metabolites and their effects.
In Fig. 2C, enrichment in GO Cellular Component (CC) is represented as a circular plot linking modulated genes to the cellular structures in which they act. Among the significantly enriched components were extracellular exosomes and endocytic vesicles, where RAB10 and ABCC4 were up-regulated (genes implicated in vesicular trafficking and metabolite export), while SLC12A2 and RCV1 were repressed. These changes indicate that the postbiotic actively regulates vesicle and exosome release, modulating intercellular communication and the transfer of pro-oncogenic signals. In the mitochondrial category, PRDX2 (peroxiredoxin-2, with antioxidant activity) was up-regulated. At the same time, ENO1 (a glycolytic enzyme essential for tumor metabolism) was repressed, pointing to a rebalancing of oxidative metabolism and a potential reduction of the glycolytic phenotype typical of cancer cells (Warburg effect). Within DNA repair complexes, RAD1 (a DNA damage sensor) was down-regulated, suggesting that the postbiotic interferes with the tumor cells’ ability to respond to genomic instability. Finally, in transcriptional regulator complexes, RFXAP and MEST were both repressed, indicating modulation of epigenetic regulation and cell differentiation programs.
Finally, Fig. 2D shows enrichment in GO Molecular Function (MF), which revealed an equally relevant profile with key functions modulated by specific genes. For transcription factor binding, CD80 (up-regulated) was associated with immune modulation through T cell activation via CTLA4/CD28 interactions. In contrast, CREB (down-regulated) is a transcription factor known to promote cell proliferation and survival, suggesting that the postbiotic exerts an inhibitory effect on proliferative programs. Regarding kinase activity and nucleotide binding, PIK3CG was strongly repressed, a central mediator of the PI3K–Akt pathway that supports tumor growth and resistance to apoptosis. Its suppression points to direct inhibition of this signaling axis. In ligand and receptor binding, VEGF and PDGFA were up-regulated, both linked to tyrosine kinase receptors. Although these are typically pro-oncogenic, their upregulation in a context of simultaneous repression of major metabolic and proliferative drivers (e.g., PIK3CG and ENO1) may reflect a compensatory feedback mechanism induced by the postbiotic. For adaptor molecular functions, GNB2 was up-regulated, while GNG11 was repressed, indicating a reorganization of G-protein–coupled receptor (GPCR) signaling cascades.
Pathway and Gene Ontology (GO) enrichment analysis of DEGs in SW480 cells treated with Bacillus mojavensis postbiotic.
Enrichment analysis revealed that the transcriptional reprogramming induced by the postbiotic involved key oncogenic and immune-related pathways (Fig. 3). KEGG pathway enrichment (Panel A) highlighted significant modulation of PI3K–Akt signaling, MAPK signaling, cytokine–cytokine receptor interaction, apoptosis, and general pathways in cancer, consistent with the repression of PIK3CG and CREB and the up-regulation of IL2, VEGF, and PDGFA. GO Biological Process enrichment (Fig. 3B) identified terms such as regulation of MAPK cascade, apoptotic signaling, cell adhesion, lymphocyte differentiation, and cellular stress response, linking postbiotic treatment to reduced proliferative capacity and enhanced immune regulation. GO Molecular Function enrichment (Fig. 3C) indicated enrichment in kinase activity, transcription factor binding, cytokine receptor binding, and growth factor binding, reflecting broad modulation of signaling and transcriptional control. Finally, GO Cellular Component enrichment (Fig. 3D) revealed significant associations with focal adhesions, exosomes, vesicles, mitochondria, and transcription regulator complexes, supported by the up-regulation of RAB10 and ABCC4 and the down-regulation of SLC12A2. Collectively, these results support the idea that the postbiotic disrupts proliferative and survival pathways while enhancing immune signaling and vesicle-mediated communication, although further studies are warranted to confirm these mechanistic interpretations.
{kind=link}