Sequence analysis of VsAPX1 gene and protein
APXs are important H2O2 scavenger antioxidant enzymes, which maintain H2O2 homeostasis under normal and stressful conditions. The cytosolic APX isoenzyme is the most studied among other isoforms, due it is highly responsive and induced under different stresses (Pandey et al. 2017; Saxena et al. 2020), as well as its role in different defense processes, and in plant homeostasis redox regulation (Caverzan et al. 2019). Guo et al. (2020), proved the role of cytosolic APX1 in maintaining redox balance to regulate cotton photosynthetic rate and yield. Also, in Arabidopsis thaliana APX1 protected chloroplast under high light intensity (Saxena et al. 2020). Therefore, the present study characterized the full-length gDNA and cDNA cytosolic ascorbate peroxidase gene from the V. sativa plant. The VsAPX1 cDNA sequence contains an in-frame start site (ATG, 62 bp) and stop site (TAG, 812 bp), these signals suggest that the VsAPX1 cDNA is full length. Furthermore, a putative polyadenylation signal site (AAATAA) was detected in the VsAPX1 sequence at a 159 bp downstream stop codon, this signal site was previously reported in other legumes (Mittler and Zilinskas 1992). Different studies have cloned and characterized cytosolic APX cDNA from different plant species such as pea (Mittler and Zilinskas 1991), sweet potato (Park et al. 2004), potato (Kawakami et al. 2002), and spinach (Ishikawa et al. 1995).
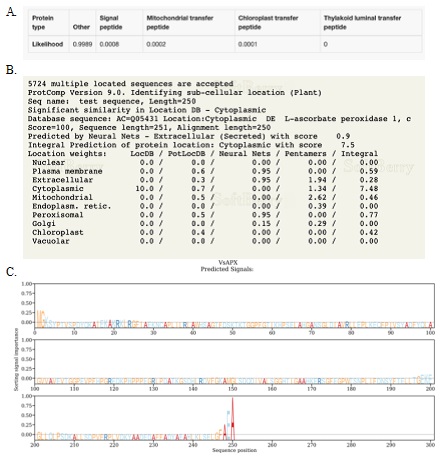
The ORF of VsAPX1 encodes 250 amino acid residues (Fig. 1), with a molecular weight of 27.1 kDa. APX isoenzymes are different in molecular weight for example, in rice chloroplast (thylakoidal APX (~ 51 kDa), stromal APX (~ 33–38 kDa)), peroxisomal APX (~ 32 kDa), and cytosolic APX (~ 27 kDa) (Teixeira et al. 2004), while in potato mitochondrial APX (~ 31 kDa) (De Leonardis et al. 2000). VsAPX1 amino acid residues lack localization signals; an N-terminal transit peptide sequence and a C-terminal transmembrane domains, according to the protein targeting analysis. This confirms the bioinformatic results that the VsAPX1 protein is cytosolic ascorbate peroxidase.
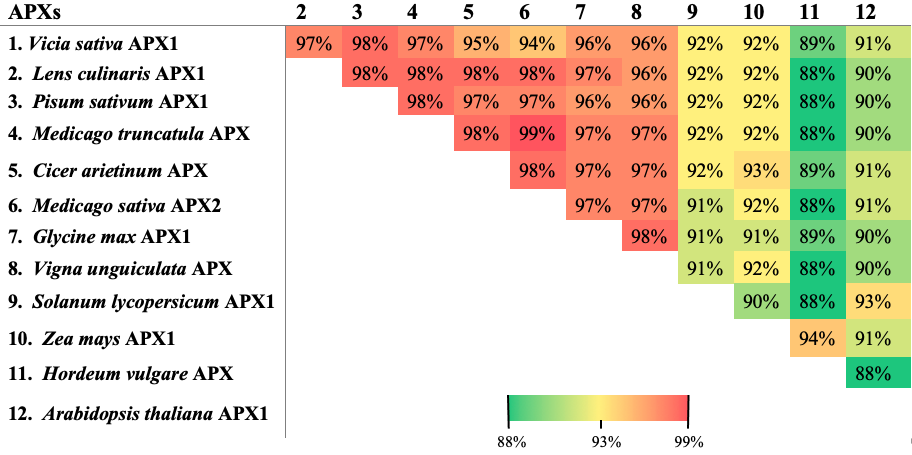
Sequence similarity analysis of VsAPX1 with closely related proteins from different plant species was investigated using BLASTp (Supplementary Fig. 2). The results showed that the deduced VsAPX1 amino acid sequence was highly similar to other cytosolic APX proteins; this could indicate that the cloned gene encodes cytosolic APX.
Ascorbate peroxidase protein, in its active form, is a dimer (containing two identical subunits), distinguished by two domains, C-terminal and N-terminal surrounded by the heme, and specified by its residues that are essential for the activity (Dąbrowska et al. 2007; Pandey et al. 2017). Ascorbate, heme (iron), and potassium ions are critical for APX activity (Jespersen et al. 1997). In the present study, the multiple sequence analysis of the VsAPX1 primary structure (Fig. 1) revealed fundamental and functional conserved motifs, domains and residues that are essential for APX protein structure and function. These include the APX active site, proximal heme-ligand motif, K+ binding site, substrate binding sites, and heme binding sites. Several studies have identified the proximal His-163 in the heme binding site and Arg-38 which are essential for heme binding. The distal His-42 and Arg-38 in the active site are responsible for the heterolytic cleavage of H2O2. The hydrogen bonding between His-163, Asp-208, and Trp-179 form the active site structure (Lazzarotto et al. 2021). The substrate ascorbate binds to the active site by four hydrogen bonds with Lys-30, Arg-172, Cys-32, and the heme moiety (Pandey et al. 2017). At the proximal domain, K+ ions bind to K+ binding sites (Thr-164, Thr-180, Asp-187) that are required for APX activity (Dąbrowska et al. 2007). Phosphorylation of Thr-59 and Thr-164 residues was reported to increase tomato APX enzyme activity, and S-nitrosylation of Cys-32 enhance APX enzyme catalytic activity (Ravi et al. 2023). APX class I is distinguished from other classes by Trp-41 and Trp-179 instead of Phe-41 and Phe-179 (Dąbrowska et al. 2007). Moreover, cytosolic APX is differentiated by Phe-175 instead of Trp-175 found in the chloroplast, and Ser-43, Phe-57, and Thr-59 are replaced by Asp-43, Asn-57, and Ser-59 in other APX isoforms (Jespersen et al. 1997). These data suggest that VsAPX1 belongs to the class I cytosolic ascorbate peroxidase.
Some studies reported a dual function for APX protein under abiotic stresses, which means functional switching; under salinity stress act as peroxidase, and under heat stress tend to be molecular chaperones. The dual function of APX resulted from structural conformation, which leads to the association of oligomer to high molecular weight (molecular chaperone) under heat stress, or dissociation of oligomers to low molecular weight (APX enzyme activity) under salinity stress.
Molecular phylogenetics deals with evolutionary relationship based on different macromolecules (DNA, RNA, and protein) (Bogusz and Whelan 2017). This study clarifies the phylogenetic relationship of VsAPX1 with APXs orthologs from other plant species, for this purpose a neighbor-joining phylogenetic tree was constructed (Fig. 4). The resulting tree showed that APX proteins were clearly separated into three groups based on their subcellular localization: cytosolic, peroxisomal, and chloroplastic, and VsAPX1 protein was clustered within the cytosolic clade which is closely related to the leguminous species Pisum sativum and Lens culinaris, indicating a close relationship between these APX proteins. This result is consistent with the phylogenetic tree generated in the study of Malambane et al. (2018). This apparent divergence between orthologs, revealed in previous studies indicating that cytosolic and peroxisomal APX isoenzymes were generated by a duplication event of a non-chloroplastic ancestral gene (Teixeira et al. 2004). According to Qu et al. (2020), results of APX genes phylogenetic tree and exons structure revealed that the ancestors of monocots and dicots underwent genome duplication. While Ozyigit et al. (2016) APX phylogenetic tree of 18 plant species showed segmental and tandem duplications in some APX genes.
Ascorbate peroxidase gene structure
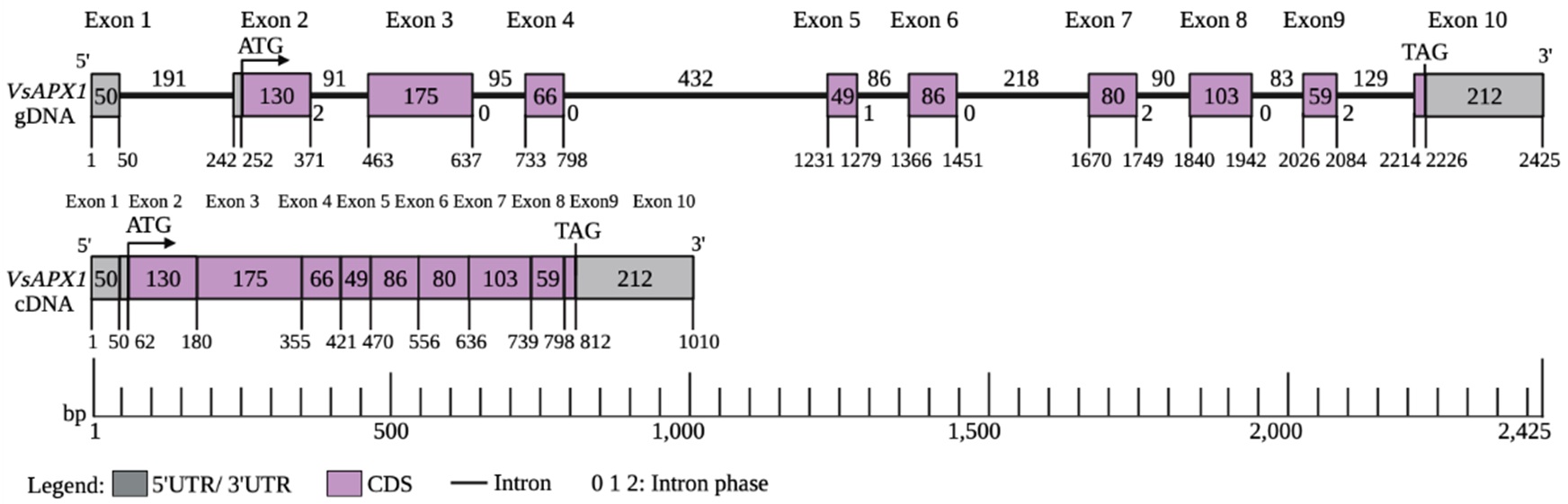
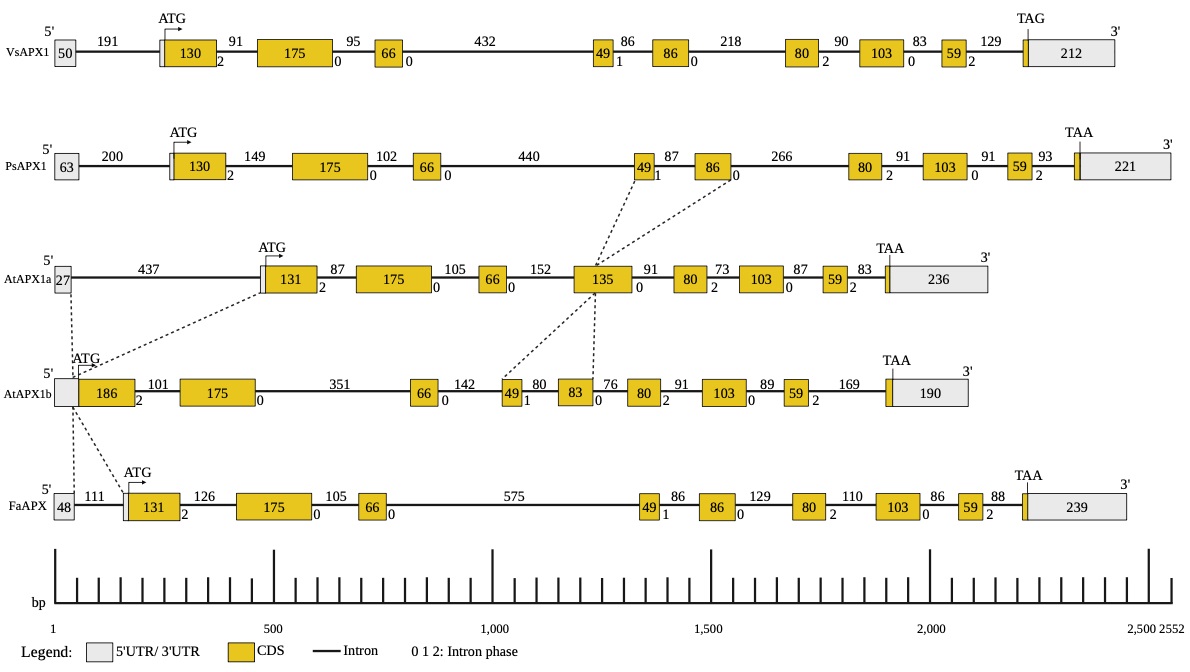
The identification of the VsAPX1 gene structure provides insights into the organization of exons and introns in this gene, which can aid in further genetic studies and manipulation of the gene. To investigate the structure of VsAPX1, the exon/intron organization was constructed and compared with other plant cytosolic APX genes. In the present study, the VsAPX1 gene contains 10 exons enterspaced by 9 introns (Supplementary Fig. 3). Comparing VsAPX1 gene structure with other plant species (Supplementary Fig. 4), the length of VsAPX1 exons of ORF 9 exons is highly identical to pea APX1 (which is the only gene available with detailed for APX in legumes) (Mittler and Zilinskas 1992), and strawberry (Kim and Chung 1998). Moreover, similar result was found in other plant APX genes such as tomato (Gadea et al. 1999) rice (Teixeira et al. 2004), maize (Liu et al. 2012; Qu et al. 2020), and wild watermelon (Malambane et al. 2018). Whereas there are differences in intron length, nucleotide sequences, while the intron number, and intron phases are conserved (Patthy 1987). This suggests a conserved cytosolic APX1 gene architecture in higher plants (Teixeira et al. 2004; Qu et al. 2020). Likewise, APX genes promoter cis-elements, exon-intron organization, and number were studied in Populus trichocarpa (Leng et al. 2021). On the other hand, Arabidopsis thaliana APX1a, APX1b had 9 exons and 8 introns (Santos et al. 1996). A major difference appears in exon5 of AtAPX1a which is separated by 86 bp intron to exon 5 and exon 6 in VsAPX1gene. The first intron was located within 5’UTR. Similar observation was made in those of other plant species except for the A. thaliana APX1b (Supplementary Fig. 4) which lacks the 5’UTR intron, the gene encoding a second family of cytosolic APX and similar observation in Dancy’ tangerine (Citrus reticulata Blanco) (Santos et al. 1996; Kunta et al. 2010). Mittler and Zilinskas (1992) detected a part of the GPEI enhancer (TGATTCAG) sequence in 5’UTR intron, which is a regulator element for glutathione transferase P that regulates gene transcription by the interaction with transcription factors and RNA II polymerase (Mittler and Zilinskas 1992). Some studies discussed the effect of 5’UTR intron, which may interact with other elements located in the promoter that regulate APX1 gene expression, in fact, the leader intron of APX20 gene in tomato increased expression of a reporter gene in leaves but it was absent in roots (Gadea et al. 1999). The gain of an intron in 5’-untranslated region and in exon number five of VsAPX1 causes exon fission but did not cause a shift in reading frame unlike the exon loss/gain that can cause a shift in reading frame (Xu et al. 2012). The intron insertion at this location may be due to the presence of proto-splice sites (G|G, and MAG|R; M: A or C, R: A or G) that increase chance of intron insertion at this position (Nguyen et al. 2006).
Expression pattern analysis of VsAPX1 gene in response to stresses
Plants have complex defense systems that protect themselves from environmental changes. These defense systems are augmented and activated by the perception of stress signaling molecules (phytohormones and ROS (H2O2)), which induce signal transduction cascades and then regulate gene expression of the stress responsive genes (Nadarajah 2020). Antioxidant enzymes scavenge the excess ROS and maintain cellular homeostasis from oxidative damage (Rajput et al. 2021). APX is one of the crucial enzymes in the AsA-GSH cycle and can scavenge excess H2O2 (Hasanuzzaman et al. 2019). Recent studies showed the up-regulation of APX gene expression under different abiotic stresses and stress-response chemicals, which could be due to the presence of stress-response cis-acting elements in the promoter that activate APX gene expression (Leng et al. 2021). These elements include phytohormone-responsive, abiotic stress-responsive, and growth and development-responsive elements. Generally, cAPX 5' regulatory region has a heat shock responsive element and anti-peroxidative element (ARE) that might aid in H2O2-scavenging (Pandey et al. 2017). The present study investigated the expression pattern of APX gene in V. sativa under phytohormone treatments (ABA, SA, and JA) and abiotic stresses (42οC and H2O2) using qRT-PCR, to understand and define the possible involvement of the VsAPX1 gene to stresses.
The increased global temperature is a critical climate-change problem, resulting in an increasing rate of evaporation and dehydration in plants and soil (Hassan et al. 2020). Heat stress resulted in the deactivation of enzymes, protein misfolding, disturbs cell metabolism, increased the fluidity of membrane lipid, and increased production and accumulation of ROS (e.g., oxidative damage) (Chiang et al. 2015). Recent studies demonstrated that some APX genes are heat inducible (Sadder et al. 2014). Under heat stress, the increasing level of cellular H2O2 acts as a signaling molecule that induces heat stress signal transduction components, including the heat stress transcription factor, which binds to the heat shock element (HSE) in the promoter of the APX gene and control its expression (Panchuk et al. 2002). This explains the quick response of APX gene to heat stress (Banerjee and Roychoudhury 2019). HSE has been determined in the APX promoter of pea (Mittler and Zilinskas 1992), and strawberry (Kim and Chung 1998). In the present study, after exposing seedlings to 42ºC, the expression of VsAPX1 scored an early increase after 2 h (Fig. 5A). Other studies found that heat stress increased APX gene expression in a variety of plant species, including pea (Mittler and Zilinskas 1992), rice (Sato et al. 2001), alfalfa (Li et al. 2013), Arabidopsis thaliana APX2 (Wang et al. 2020b), and sweet potato APX1 (Park et al. 2004). Moreover, heat stress increased the activity of the cucumber cAPX enzyme (Song et al. 2005). The overexpression of pea cAPX enhanced heat tolerance in transgenic tomato (Wang et al. 2006). Furthermore, the heat tolerance of Arabidopsis thaliana was increased by the overexpression of cabbage APX gene (Chiang et al. 2015).
H2O2 is an important non-radical ROS, generated from normal cellular metabolism as a harmful by-product that cause damage and inactivation for cellular components (Das and Roychoudhury 2014; Mittler 2017). At basal levels, H2O2 acts as a regulatory signal for different physiological processes including photosynthesis, photorespiration, stomatal closure, growth, cell cycle, development, and senescence (Hasanuzzaman et al. 2020). Furthermore, under stress conditions, H2O2 acts as a signaling transduction molecule, due to being highly stable and diffusible, can pass through the plasma membrane via aquaporins and diffuse from different organelles and transport the signal to the nucleus through redox reactions, integrated with the MAPK pathway (Singh et al. 2019), which involved in the regulation of nuclear gene expression of many transcription factors, and upregulation of antioxidative enzymes (Song et al. 2005). The current study reports a gradual decline in the VsAPX1 gene expression over time under foliar application of H2O2, this may be due to the high concentration of H2O2 (Fig. 5B). Moskova et al. (2009) reported that exogenous application of H2O2 decreased APX enzyme activity in pea. In contrast to other studies, the exogenous application of H2O2 increased gene expression in sweet potato (Park et al. 2004), and cucumber cAPX enzyme activity (Song et al. 2005). In addition, H2O2 treatment was reported to induce APX gene expression in cultured soybean cells (Lee et al. 1999). This is possibly due to the presence of an Antiperoxidative element (ARE) in the APX promoter, which could be involved in the regulation of the APX gene and responsible for the H2O2-induced response (Mittler and Zilinskas 1992; Lee et al. 1999).
Expression Analysis of VsAPX1 Gene in Response to Phytohormones
Plant bioregulators are chemicals found in small quantities that have a large influence on regulating plant growth, development, and yield under normal and stressful conditions (Alhaithloul et al. 2020; Hasanuzzaman et al. 2021). Whereas, under unfavorable environmental conditions, plant hormones (ABA, JA, SA, and ethylene) play a role in plant adaptation and regulate the balance between plant development and stress responses (Wang et al. 2020a). Furthermore, the exogenous application of phytohormones at a suitable dose enhanced plant tolerance to abiotic stress conditions (Awan et al. 2021).
ABA acts as a stress-responsive hormone that plays a crucial role in plant adaptation (Gong et al. 2020). Its biosynthesis increases under stress conditions and regulates the expression of different stress-responsive genes and protective proteins like antioxidant enzymes, late embryogenesis abundant proteins, and dehydrins (Wani et al. 2016). Therefore, exogenous ABA treatment was reported to increase the expression and activity of ascorbate peroxidase in different plant species, which is possibly due to the presence of APX promoter cis-elements antioxidant responsive element and ABA responsive element (Saxena et al. 2020). In this study, the foliar application of ABA markedly increased the VsAPX1 expression after 2 h, this indicates that ABA has a signaling effect on VsAPX1 expression. Similar observations were reported after ABA treatment in pea (Mittler and Zilinskas 1992), maize (Liu et al. 2012), and sweet potato (Park et al. 2004).
SA acts as a growth regulating and protector molecule that improves crop plant tolerance under stress conditions (Alhaithloul et al. 2020), and induces the gene expression of antioxidants, HSPs, chaperones, and genes involved in the biosynthesis of secondary metabolites (Wani et al. 2016). Foliar application of SA under abiotic stresses enhances the activity of antioxidant enzymes and reduces oxidative stress impacts such as lipid peroxidation and membrane injury (Pourghasemian et al. 2020). The current study showed a slight rise in the VsAPX1 transcript level after 2 h of foliar SA treatment. Some studies reported that SA application increased endogenous H2O2 accumulation and increased the expression and activity of antioxidant enzymes (Dikilitas et al. 2020). The SA application increases the transcript level of sweet potato cytosolic ascorbate peroxidase (Park et al. 2004), and pea APX enzyme activity (Yadu et al. 2017).
JA aids in plant reproduction, development, tendril coiling, fruit ripening, chlorosis, allelopathy, production of secondary metabolites, flower and seed development, wounding and herbivory, and leaf senescence (Awan et al. 2021). Whereas, under a stressful environment, JA act as a signaling molecule and stress-responsive hormone (Alisofi et al. 2020). JA was reported to interact with other plant hormones, transcription factors, and enhanced the expression of JA-associated genes and stress-responsive genes and increased the activity of the antioxidant defense system (Wang et al. 2020a). VsAPX1 transcript level was highly accumulated after 4 h of JA application. Similar observation was recorded in rice OsAPX1 (Wang et al. 2015), and sweet potato (Park et al. 2004). The foliar application of JA alone or in combination with salinity on bitter melon seedlings decreased the activity of APX enzyme (Alisofi et al. 2020).
{kind=link}
{kind=link}
{kind=link}
{kind=link}