Compositional analysis of C. vulgaris
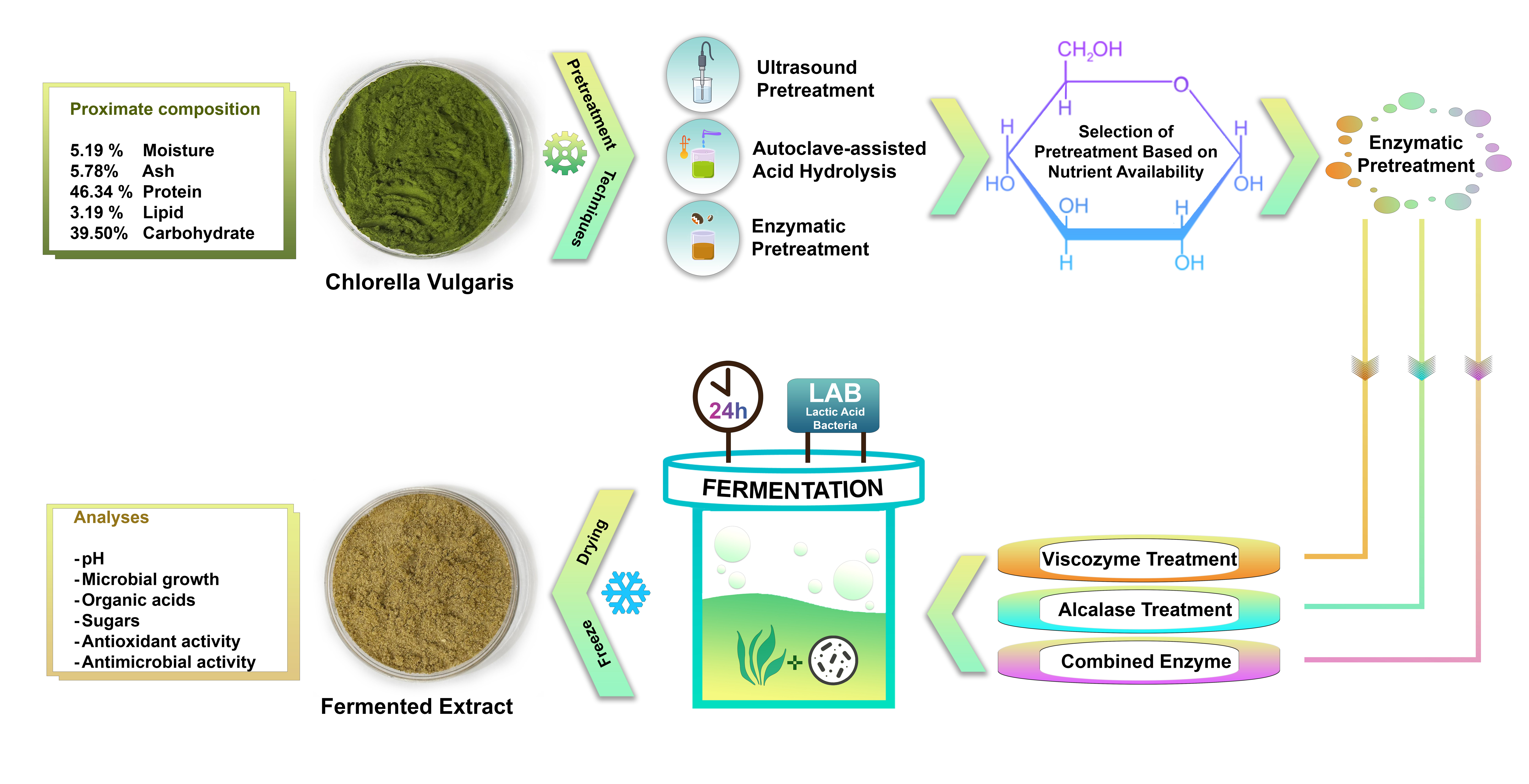
In the first part of this study, the proximate composition of C. vulgaris biomass was analysed, with the results presented in Fig. 1. As shown, proteins (~ 46%) and carbohydrates (~ 40%) were the primary components of the studied biomass. In comparison, lipid (~ 3% ) and ash contents (~ 6%) were comparatively lower (Fig. 1). These findings are consistent with a recent study, which reported ash, lipid, protein, and carbohydrate contents of approximately 9%, 0.5%, 40%, and 43%, respectively (Damayanti et al. 2025). Furthermore, previous reviews reported that the total protein content in mature C. vulgaris ranges from 42% to 58% of the biomass dry weight (Safi et al. 2014; Ijaola et al. 2024). Although carbohydrate levels are typically lower, Chlorella is known to accumulate high starch contents, particularly under nitrogen-limiting conditions, reaching a total carbohydrate content up to 55% of its dry weight (Brányiková et al. 2011). The observed lipid content in this study is lower than most reported values in the literature, ranging from 5% to 58% with higher content typically obtained under mixotrophic cultivation (Wang et al. 2024). Nevertheless, the biochemical and nutritional profile of C. vulgaris is highly dependent on cultivation conditions and the strain used (Hiltunen et al. 2026).
Evaluation of various pretreatment strategies
The first method used to disrupt the cell wall of C. vulgaris was ultrasound treatment, applied in distilled water or sulfuric acid solutions at varying concentrations (Fig. 2). Ultrasound-assisted extraction is a green technology used to recover bioactive compounds from algae with reduced solvent and energy input, while preserving molecular integrity. Ultrasound induces cavitation in liquid media, and the collapse of these microbubbles generates localised mechanical forces that disrupt the cell walls of microalgae, facilitating the release of intracellular compounds (Lee et al. 2017). As a technique, it has been widely applied to extract polysaccharides, pigments, and phenolics from various algal species (Carreira-Casais et al. 2021). The highest release of reducing sugars (~ 9 mg g DW− 1) was achieved with 2% sulfuric acid (Fig. 2A), while the maximum protein content was observed with 4% acid. However, no statistically significant difference was observed compared with 2% acid (Fig. 2B). As shown, even at the highest yield achieved with this method, the carbohydrate conversion is relatively low, reaching 2.2%. This result closely agrees with a previous study utilising ultrasonic-assisted acid hydrolysis for Chlorella, which reported a carbohydrate conversion efficiency of 3.3%. The authors concluded that the application of ultrasonic treatments as the only method for releasing and depolymerising carbohydrates is insufficient (Nasirpour et al. 2023). Although ultrasound treatments are a useful pretreatment method in other applications, they may not achieve sufficient conversion of complex carbohydrates into simple sugars, which are essential for subsequent fermentation.
In the following step, hydrothermal hydrolysis was conducted at 121°C for 30 minutes using sulfuric acid concentrations of 1%, 2%, and 4% (v/v), with distilled water serving as the control. The results, summarised in Fig. 3, indicate that hydrolysis with 2% sulfuric acid yielded the highest amount of reducing sugars, reaching 102.4 mg g DW− 1 (Fig. 3A). On the other hand, the lowest yield was observed for hydrothermal treatment with water as solvent, reaching a conversion efficiency of only 0.9%. A similar observation was also reported in a study on Chlorella, where the conversion efficiencies at 30 and 120 minutes of heating in the hydrothermal method were similarly low, at 0.5% and 1%, respectively (Nasirpour et al. 2023). Protein recovery was significantly higher in samples treated with 2% and 4% acid than in those treated with lower concentrations, although the difference between the two highest concentrations was not statistically significant (Fig. 3B).
Regardless of the technique used, the addition of sulfuric acid resulted in a substantially higher release of reducing sugars and proteins (Figs. 2 and 3). Dilute sulfuric acid is commonly employed for lignocellulosic biomass pretreatment due to its rapid reaction kinetics and enhanced cellulose hydrolysis efficiency (Esteghlalian et al. 1997; Sun and Cheng 2002). In practically all cases, a 2% sulfuric acid addition resulted in the highest recoveries. This observation aligns with a report by de Carvalho Silvello et al. (2023), who investigated the effect of 1%, 2.5%, and 4% sulfuric acid concentrations on the release of sugar from biomass. The authors indicated that 2.5% sulfuric acid yielded the highest sugar release, while increasing the concentration to 4% did not result in a significant improvement (de Carvalho Silvello et al. 2023). Moreover, in another study, the effects of 2% and 5.36% sulfuric acid at 120°C for 30 min on sugar release were compared, with the authors reporting that the highest yield (247 mg/ gDW) was achieved with the 2% acid concentration (Abou El-Souod et al. 2021). Nevertheless, it is worth noting that a previous study comparing five acids for C. vulgaris hydrolysis found that while hydrochloric, sulfuric, and nitric acids were more effective than peracetic and phosphoric acids, hydrochloric acid yielded the best overall results (Park et al. 2016). High recoveries (> 80%) have also been reported by other studies utilising hydrochloric acid for the pretreatment of Chlorella biomass towards bioethanol production (Kim et al. 2011; Zhou et al. 2011). Nonetheless, during acid hydrolysis, especially at elevated temperatures, carbohydrate degradation often leads to the formation of furanic compounds, such as hydroxymethylfurfural and furfural, which are known to inhibit microbial activity and reduce fermentation efficiency (Labbafi et al. 2025).
Considering these factors, the next step was to apply three enzymatic treatments to disrupt the cell wall matrix and enhance the release of intracellular compounds. Enzymatic hydrolysis has been proposed as an effective method to disrupt the cell wall of C. vulgaris, thereby facilitating the release of valuable intracellular components, including proteins, carbohydrates, and bioactive compounds (Rahman et al. 2022). Towards this, Viscozyme, a multi-enzyme complex with carbohydrase activity, was used to break down polysaccharides, while alcalase, a protease, targeted protein structures. Additionally, a combined treatment using both enzymes was performed to maximise cell wall degradation and improve nutrient availability for downstream processes. As shown in Fig. 4A, enzymatic hydrolysis, particularly with viscozyme alone and in combination, significantly increased the release of reducing sugars, yielding 139.61 and 151.18 mg g DW− 1, respectively, compared to 4.34 mg g DW− 1 in the control samples. These findings are in close agreement with a previous report, which reported that the highest soluble carbohydrate hydrolysis (160 mg g DW− 1) was obtained with the carbohydrolase (V and V + A) after 2 h (Mahdy et al. 2014). On the other hand, alcalase treatment resulted in the highest protein release, reaching 14.4 mg g DW− 1, corresponding to an extraction efficiency of 3.1%. Similar recovery values were reported in a previous study, where the enzymatic protein extraction process yielded an average efficiency of 3.80% (Yaghoubzadeh and Safari 2025).
Overall, among the various tested techniques, ultrasound resulted in the lowest recovery of fermentable sugars and protein, followed by the hydrothermal treatments. The low efficiency of ultrasound has also been reported in previous studies. For instance, de Carvalho et al. evaluated alkaline, acid, ultrasonic, and supercritical treatments and found that ultrasound in water (10 min, 200 W, 20 kHz) yielded less than 1 g/L of glucose, one of the lowest values among the tested methods (de Carvalho Silvello et al., 2023). Similarly, Paterson et al. applied ultrasound after freeze–thaw cycles, but microscopy revealed insufficient cell wall disruption, highlighting the need for more effective strategies (Paterson et al. 2024). Hydrothermal treatment also did not result in satisfactory results in this study. However, it should be noted that a previous report indicated that although the hydrothermal or ultrasonic treatment alone may be insufficient, their combination results in substantially higher recoveries of fermentable sugars (Nasirpour et al. 2023). The effectiveness of combinatorial approaches has also been suggested in other reports, where alkali and enzyme hydrolysis produced the highest reducing sugar yield (Effiom et al. 2023).
In this study, enzymatic treatments resulted in substantially higher recoveries of fermentable sugars, especially as compared to the other two methods. The efficiency of cellulases and alcalases in converting the complex carbohydrates of Chlorella into fermentable sugars has been demonstrated for biohydrogen (Liu et al. 2021; Sriyod et al. 2021) and bioethanol (Kim et al. 2014). In fact, a recent review concluded that compared to other pretreatment methods, enzymatic hydrolysis offers advantages for bioethanol production due to its low cost, minimal environmental impact, and higher ethanol yields, as it also operates under mild conditions, requiring less energy and no harsh chemicals, thereby avoiding the formation of fermentation inhibitors (Kusmiyati et al. 2023). Besides bioenergy production, enzymatic hydrolysis has also been used as a strategy to increase the functional properties of Chlorella hydrolysates (Yin et al. 2010; Gharehbeglou et al. 2024). An additional benefit of this approach is the absence of fermentation inhibitors. Considering all these factors, enzyme-assisted hydrolysis was selected as the optimal strategy prior to LAB fermentation, as discussed in the following sections.
Fermentation of C. vulgaris by lactic acid bacteria
Microbial growth and acification during LAB fermentation
In this part of the study, C. vulgaris biomass underwent enzymatic hydrolysis prior to fermentation by LAB. To evaluate the effectiveness of the fermentation process and the capacity of pretreated biomass to support microbial activity, the growth of inoculated LAB strains was monitored and expressed as log CFU mL⁻¹. Concurrently, pH was monitored to assess acidification of the fermentation medium. The results of these parts are summarised in Fig. 5. Before inoculation, the cell densities of L. plantarum and L. brevis were standardised to approximately 6 log CFU mL⁻¹ to ensure consistent initial microbial loads across all fermentation trials. As depicted in Fig. 5, both strains exhibited substantial growth after 24 hours of fermentation. In samples fermented with L. plantarum, the highest growth of 10.12 log CFU mL⁻¹, corresponding to a Δ Log CFU mL⁻¹ of 4.1, was observed for the sample treated with Viscozyme (FVPL, Fig. 5A). The other two treatments, FAPL and FVAPL, also showed a significant increase in cell density with Δ Log CFU mL⁻¹ of 3.6 and 4.0, respectively, thereby overall indicating robust proliferation under the given conditions (Fig. 5A).
Similar trends were observed in samples inoculated with L. Brevis, where all treatments supported effective growth. The Viscozyme-treated samples (FVBR) reached the highest cell density of 9.76 log CFU mL⁻¹, with Δ Log CFU mL⁻¹ of 4.1 (Fig. 5C). The other treatments also showed strong LAB proliferation, with increases of ~ 3.9 Log units.
In addition to microbial growth, acidification of the medium was used as an indicator of fermentation efficiency. As illustrated in Fig. 5B and 5D, both LAB strains demonstrated significant acidification capacity within 24 hours of fermentation. The lowest pH values were recorded in the Viscozyme-treated samples, reaching 3.6 for L. plantarum (Fig. 5B) and 4.2 for L. brevis (Fig. 5D). In contrast, the lowest acidification for both strains was observed in the alcalase-treated samples.
These findings align with previous studies that have reported the beneficial effects of Chlorella species on the viability and growth of probiotic strains such as Lactobacillus and Bifidobacterium. In most cases, Chlorella biomass was incorporated either into fermented semi-solid dairy matrices such as cheese and yoghurt (Kovaleski and Ventura 2025) or into freeze-dried probiotic formulations (Meireles Mafaldo et al. 2022; Fortuin et al. 2026). For instance, Ścieszka and Klewicka demonstrated that supplementing MRS medium with varying concentrations of C. vulgaris biomass accelerated the growth of four L. brevis strains, shortening their logarithmic growth phase (Ścieszka and Klewicka 2020). Moreover, another study reported that C. vulgaris increased the survival of Lactobacillus spp. in bile salts. They noted that the protective effect of algae at low pH is strain-dependent, concluding that the biomass of this microalga could be used in fermented dietary products (Sylwia and Elżbieta 2020).
However, only a limited number of studies have explored the use of C. vulgaris biomass as a sole carbon source for LAB fermentation. In a recent study, Niocolloti et al. investigated the fermentation of four C. vulgaris biomasses with varying carbohydrate content using 5 LAB strains, including L. plantarum 4932. Their results showed that all biomasses fermented with L. plantarum reached pH values below 4.5 after 72 hours of fermentation, although microbial growth was modest, reaching a maximum of 2.1 (Nicolotti et al. 2025). Moreover, another study reported co-fermentation of Aphanizomenon flos-aquae and C. vulgaris biomasses with L. plantarum and S. cerevisiae; however, the authors do not report either cell growth or pH values (Tomassi et al. 2025). Taken together, the results of the present study highlight the importance of enzymatic pretreatment in enhancing the fermentability of C. vulgaris biomass. Compared to previous reports, the observed microbial proliferation and acidification suggest that enzymatic hydrolysis significantly improves the bioavailability of fermentable substrates, thereby supporting more efficient LAB growth and metabolic activity.
Analysis of sugars and organic acids
In parallel with the growth experiments, the reducing sugar content and organic acid production were monitored, with results summarised in Fig. 6. Generally, L. plantarum exhibits facultative heterofermentative metabolism, meaning its fermentation products can differ depending on the environmental conditions (Siezen and van Hylckama Vlieg 2011). Most LAB thrive in anaerobic environments, where they convert pyruvate produced via the Embden-Meyerhof-Parnas pathway into lactate. On the other hand, L. brevis is an obligate heterofermentative LAB, as it consistently produces a mixture of lactic acid, carbon dioxide, and ethanol or acetic acid during fermentation, regardless of culture conditions. Unlike facultative fermenting bacteria, it relies on the phosphoketolase pathway to metabolise sugars. A significant reduction in reducing sugar concentration was observed after 24 h of fermentation (Supplementary Material Fig. S1). As compared to alcalase-treated samples, sugar consumption was higher in viscozyme-treated and combined-treated samples. Chromatographic analysis confirmed that the hydrolysates primarily contained glucose, fructose, and xylose, along with maltose (Fig. 6 and Supplementary Material Fig. S2). Due to chromatographic limitations, fructose and xylose were not fully resolved. These findings are consistent with previous reports on the carbohydrate composition and enzymatic hydrolysis of C. vulgaris, which typically release glucose, fructose, xylose, and maltose (Safi et al. 2014; Mahdy et al. 2014; El-Naggar et al. 2020). Both LAB strains exhibited similar overall sugar consumption, except in Alcalase-treated samples, where L. plantarum consumed more sugars than L. brevis. Xylose utilisation was greater in L. brevis, aligning with its metabolic capacity to ferment pentoses via the phosphoketolase pathway (Zhang et al. 2016). This pathway also explains the small amounts of acetic acid detected in samples fermented by L. brevis (Supplementary Fig. S3).
Correspondingly, the production of organic acid is presented in Fig. 7. The sample treated with combined enzyme and fermented with L. plantarum exhibited the highest lactic acid production among all tested conditions, at 17.35 g L-1, followed by the sample treated with viscozyme alone, at 12.20 g L− 1. These findings align with the measured reducing sugar, as viscozyme- and combined-treated samples showed higher sugar release from the matrix, confirming the increased conversion to lactic acid. Besides, a small concentration of acetic acid production was observed in the samples, including those treated with alcalase and combined enzyme treatments, and fermented by L. brevis, with values of 1.03 and 1.10 g L− 1, respectively (Supplementary Materials Fig. S3).
The enhanced fermentation performance observed in Viscozyme and combined enzyme treatments further highlights the importance of cell wall disruption in microalgae. Several reports on enzymatic pretreatment of Chlorella species for biofuel and bioproduct generation have shown similar benefits, including increased sugar release and improved microbial activity (Córdova et al. 2019; Yang et al. 2023). In a previous study, the authors explored fermentation strategies for lactic acid production with immobilised L. plantarum 23 using C. vulgaris hydrolysate as feedstock. They tested various aeration conditions to determine the oxygen requirements for this strain to be homolactic (Chen et al. 2020). The study concluded that although lactic acid’s yield and productivity were not substantially different under the different oxygen availability, the strain was homolactic only under anaerobic conditions (Chen et al. 2020).
Nevertheless, the ability of C. vulgaris to serve as a sole carbon source for LAB fermentation, especially when pretreated enzymatically, could open new avenues for sustainable bioprocessing. This strategy could also be considered particularly relevant for developing functional foods and probiotic formulations, as highlighted by recent work on engineered C. vulgaris strains with enhanced carbohydrate content (Saha et al. 2024).
Antioxidant activity of fermented C. Vulgaris
The antioxidant capacity of C. vulgaris biomass and its extracts is primarily associated with a diverse range of bioactive compounds, including chlorophyll, carotenoids (such as lutein and β-carotene), polysaccharides, and polyphenols (Mendes et al. 2024). Although fermentation has been proposed as a strategy to enhance the antioxidant activity of microalgae, data on its impact on C. vulgaris and its extracts remain limited. In this study, the in vitro antioxidant capacity of fermented C. vulgaris was assessed using four complementary assays: CUPRAC, total phenolic content (TPC), ABTS, and DPPH radical scavenging tests (Fig. 8 and Supplementary Material Fig. S4). Enzyme-treated but non-fermented samples served as controls (0h in all figures). Across all assays, combined enzyme treatment yielded the highest antioxidant capacity, with values of 8.64 mg Trolox g DW⁻¹ (CUPRAC), 9.67 mg GAE g DW⁻¹ in the TPC assay (Fig. 8). Similar observations with 19.46 mg Trolox g DW⁻¹ in the ABTS assay, and 1.45 mg Trolox g DW⁻¹ in the DPPH assay were recorded (Supplementary Material Fig. S4). However, most differences between fermented and enzyme-treated samples were not statistically significant.
When a single enzyme pretreatment was applied, the effects of fermentation became more evident, with fermented samples generally exhibiting higher antioxidant activity than unfermented ones. This observation aligns with recent findings by Tomassi et al., who reported that fermentation of C. vulgaris using LAB and yeast significantly improved its antioxidant profile. Specifically, TPC nearly doubled from 2.4 mg GAE g DW⁻¹ at the onset of fermentation to 4.8 mg GAE g DW⁻¹ after 24 hours, with similar trends observed for DPPH and ORAC assays (Tomassi et al. 2025).
The increase in antioxidant capacity observed in this study can be attributed to several mechanisms. Enzymatic pretreatment with cellulases (e.g., Viscozyme) facilitates the release of intracellular antioxidants, including phenolics, carotenoids, chlorophylls, and peptides (Sanjeewa et al. 2023; Badrunanto et al. 2025). Moreover, although proteases can contribute to cell wall disruption, the observed increase in antioxidant capacity is attributed to the generation of bioactive peptides with antioxidant properties. These observations are further supported by previous studies on other substrates that have shown similar effects with proteases and cellulases. For instance, Viscozyme was utilised to obtain a high-yield bilberry extract with enhanced antioxidant capacity (Syrpas et al. 2021). Alcalase was used to generate bioactive peptides from mesopelagic fish protein hydrolysates (Hayes et al. 2024). In a study involving lentil proteins, the highest content of phenolic compounds (3.8 mg GAE/g) was achieved through sequential hydrolysis using Alcalase and Flavourzyme (Rezvankhah et al. 2021) .
Additionally, studies suggest that fermentation enhances antioxidant capacity by breaking down complex or bound antioxidant compounds into more bioavailable and active forms, and by generating new bioactive molecules such as peptides and vitamins (Verni et al. 2019). These effects depend on the specific metabolic activities of the microbial strains used. Overall, these findings demonstrate that combining enzymatic pretreatment with microbial fermentation is an effective strategy for improving the biofunctional value of C. vulgaris. From an application perspective, such improvements expand the potential of microalgal biomass as a natural antioxidant source for food fortification, nutraceuticals, and cosmetic formulation, areas where stability and natural origin are increasingly prioritised.
Antimicrobial activity of fermented C. vulgaris
Alternative antimicrobial products have gained increasing attention as a response to rising drug resistance and environmental concerns. As a result, the antimicrobial potential of microalgae as a source of sustainable and effective bioactive compounds for diverse applications has been explored (Xia et al. 2021). In fact, extracts from C. vulgaris exhibit antimicrobial properties (Kitada et al. 2009; Fassi Fihri et al. 2024), making this species a promising candidate for the development of functional ingredients for food and pharmaceutical use (Yuan et al. 2020). To assess this potential, the antimicrobial activity of fermentation supernatants was evaluated against five bacterial strains (Table 1). Among the tested samples, only those treated with Viscozyme and fermented with L. plantarum displayed antimicrobial effects.
In contrast, neither the unfermented samples nor those fermented with L. brevis inhibited any strain. The strongest activity was observed in the Viscozyme-treated sample (FVPL), which inhibited all strains except P. aeruginosa, with zones ranging from 8.6 mm (S. aureus) to 12.4 mm (M. luteus). Moderate inhibition was noted against E. coli (Table 1). Enzymatic hydrolysis using alcalase did not enhance antimicrobial properties, as no inhibitory effects were detected in alcalase-treated samples.
Table 1
Antimicrobial activity of enzyme-treated and fermented C. vulgaris by L. plantarum and L. brevis
| | Inhibition Zone, mm |
| S. Aureus | B. Subtilis | M. Luteus | E. Coli | P. aeruginosa |
| UPL | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| UBR | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 |
| VPL | 8.64 ± 0.01 | 10.37 ± 0.06 | 12.38 ± 0.13 | 9.64 ± 0.03 | 0.00 ± 0.00 |
| VAPL | 7.79 ± 0.13 | 7.90 ± 0.05 | 9.54 ± 0.37 | 0.00 ± 0.00 | 0.00 ± 0.00 |
Results represent mean ± SD. (UPL and UBR: untreated biomass and fermented with L. plantarum and L. brevis, respectively; VPL: viscozyme treated and fermented by L. plantarum; VAPL: combined enzyme treated and fermented by L. plantarum).
These findings are consistent with previous work on macroalgae, in which alcalase treatment failed to confer antimicrobial activity across the species tested (K. Habeebullah et al. 2020). The activity observed in Viscozyme-treated samples likely reflects efficient carbohydrate release, supporting the higher metabolic activity of L. plantarum. Generally, the antimicrobial effects of LAB-fermented products are linked to organic acid production, peptide or proteinaceous bacteriocins, and other small molecules (Ibrahim et al. 2021). Among these, lactic acid is notable for lowering pH and permeabilising the membranes of Gram-negative bacteria, such as E. coli, thereby enhancing the action of other antimicrobial compounds (Alakomi et al. 2000). However, fermentate activity typically cannot be attributed to a single molecule (Figueroa et al. 2024). In fact, the absence of activity in L. brevis-fermented samples suggests that the antimicrobial effect of L. plantarum is likely due to bacteriocin production. Many L. plantarum strains synthesise plantaricins, bacteriocins of interest for their diverse applications, including food biopreservation, mitigation of irritable bowel syndrome symptoms, and protection against urinary tract infections (Abdulhussain Kareem and Razavi 2020). L. plantarum strains that produce plantaricin are valued for their gut-friendly characteristics, broad carbohydrate utilisation, pleasant sensory profile, and probiotic properties, making them highly suitable for applications in the food industry (Goel and Halami 2023).
{kind=link}