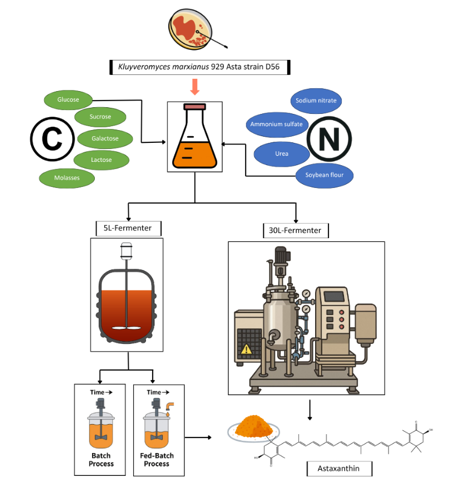
The screening of carbon sources and nitrogen sources on recombinant astaxanthin production
Several carbon sources were evaluated at a concentration of 40 g/L in flask cultures to support the growth of recombinant K. marxianus 929 Asta strain D56 and enhance astaxanthin accumulation. The tested carbon sources included glucose, sucrose, lactose, galactose, and molasses (Fig. 1). Among these, glucose supported the highest biomass growth, while galactose resulted in the greatest astaxanthin accumulation per unit of biomass. This outcome aligns with the design of the recombinant system, which utilizes galactose-inducible promoters (LAC4) and glucose-inducible promoters (Adh1) to activate the astaxanthin biosynthetic pathway. Even so, the enhancement of using galactose to be the carbon source didn’t create a significant difference on astaxanthin content as compared to using glucose.
The commercial application of galactose is severely limited due to its significantly higher cost—approximately twice that of glucose. From an industrial perspective, the trade-off between yield and production cost becomes a critical factor. Although galactose can trigger stronger pathway induction, the overall astaxanthin yield per cultivation volume may still be lower due to reduced biomass formation compared to glucose-based cultures. In contrast, glucose, despite its lower induction efficiency, supports much higher biomass production and is economically advantageous. This higher biomass compensates for the lower per-cell astaxanthin content, ultimately resulting in a competitive volumetric astaxanthin yield.
Taken together, considering the balance of pathway activation, biomass generation, and cost efficiency, glucose remains the most practical and commercially viable carbon source for large-scale astaxanthin production using recombinant K. marxianus. Further optimization of glucose-based fermentation, such as process control and feeding strategies, may help close the gap in induction efficiency while maximizing overall productivity.
As discussed earlier, glucose was identified as the most suitable carbon source for astaxanthin production in recombinant K. marxianus 929 Asta strain D56, primarily due to its cost-effectiveness. To further examine the impact of glucose concentration on astaxanthin biosynthesis, flask experiments were conducted using initial glucose concentrations of 20, 40, and 60 g/L. As illustrated in Fig. 2, biomass growth increased proportionally with higher glucose concentrations, indicating that glucose effectively supports cell proliferation. However, astaxanthin content (expressed as mg per g of biomass) exhibited an inverse relationship, decreasing as the glucose concentration increased. Conclusively, the results shown in Fig. 2 suggested 40 g/L of glucose can create the best astaxanthin production in the flask trials.
This phenomenon suggests that astaxanthin accumulation is influenced by factors beyond carbon availability, possibly linked to metabolic or environmental stresses. Specifically, a noticeable drop in final pH was observed with increasing initial glucose levels, with pH declining to around 6.0 at 60 g/L glucose. It is plausible that this acidification creates suboptimal conditions for the enzymes involved in the astaxanthin biosynthetic pathway, thereby suppressing its production despite robust biomass growth. These results highlight the delicate balance between substrate concentration, cellular growth, and metabolic output, emphasizing that optimizing pH and other fermentation conditions may be crucial to maximizing astaxanthin yield.
In addition to the carbon source, the choice of nitrogen source plays a crucial role in the cultivation of K. marxianus 929 Asta strain D56 for astaxanthin production. Peptone is commonly used as a nitrogen source in microbial cultivation due to its high nutritional value. However, its relatively high cost limits its industrial application. To explore more cost-effective alternatives, several organic and inorganic nitrogen sources—sodium nitrate (NaNO₃), ammonium sulfate ((NH₄)₂SO₄), urea, and soybean meal—were tested as partial substitutes. In these experiments, 50% of the peptone was replaced by each alternative nitrogen source, while glucose was maintained at 40 g/L.
The results of these flask trials are presented in Fig. 3. As illustrated, pure peptone remained the most effective nitrogen source for supporting the growth and astaxanthin production of engineered K. marxianus 929 Asta strain D56. Although some alternative nitrogen sources showed moderate performance, none matched the yield obtained with 100% peptone. This suggests that while partial replacement is feasible, peptone still provides the optimal nutritional balance for maximizing recombinant astaxanthin synthesis. Building on previous findings that identified glucose as the preferred carbon source, further experiments were conducted to optimize its concentration in a controlled fermentation setting. Initial flask trials indicated that while increasing glucose concentrations from 20 g/L to 60 g/L enhanced biomass production, high glucose levels led to significant pH drops, potentially inhibiting K. maximum growth.
Batch and Fed-batch operation for Astaxanthin production in a 5L-fermenter
To further evaluate the effects of initial glucose on astaxanthin production in recombinant K. marxianus 929 Asta strain D56, cultivation was carried out in a pH-controlled 5 L stirred-tank fermenter using initial glucose concentrations of 40, 60, and 80 g/L. As shown in Fig. 4, biomass increased with rising glucose levels, reaching a maximum of 16.1 g/L at 80 g/L glucose. The effect of initial glucose concentration on both biomass and astaxanthin content is summarized in Table 1. The astaxanthin content increased from 2.24 mg/g at 40 g/L glucose to 2.55 mg/g at 80 g/L glucose, demonstrating that higher glucose levels not only promote cell growth but also slightly enhance astaxanthin accumulation. A higher biomass obtained at high initial glucose resulted in greater total astaxanthin production, with the highest yield of 41.06 mg/L observed at 80 g/L glucose. Notably, glucose was completely consumed within 24 hours in all batches, indicating that carbon source depletion may limit biomass growth and, consequently, total astaxanthin output. To address this limitation, a glucose fed-batch strategy was implemented to enhance biomass accumulation and improve overall productivity.
Table 1
Effects of glucose concentrations of 40, 60 and 80 g/L in a 5L agitation fermenter
Glucose concentration (g/L) | Max. biomass (g/L) | Astaxanthin content (mg/g) | Astaxanthin Yield (mg/L) | Astaxanthin Productivity (mg/L hr) |
|---|
40 g/L | 13.51 | 2.24 | 30.26 | 0.42 |
60 g/L | 14.77 | 2.49 | 36.78 | 0.51 |
80 g/L | 16.10 | 2.55 | 41.06 | 0.56 |
The total glucose concentration was set up at 80 g/L for the batch and for the fed-batch operations of engineered K. marxianus 929 Asta strain D56 for astaxanthin production. Two different fed-batch strategies of glucose feeding were performed in this study for the comparison with batch operation. All the operations are based on the total glucose concentration of 80 g/L. The first fed-batch operation used the initial glucose of 60 g/L and fed another 20 g/L of glucose after 24 hours cultivation (marked as 60 + 20). The 2nd fed-batch operation used 40 g/L of initial glucose and fed 20 g/L and another 20 g/L at the 24th and 48th hours respectively (marked as 40 + 20 + 20). The results were shown in Fig. 5 and Table 2, which indicated the batch operation at 80 g/L can have the best performance of biomass of 16.1 g/L as compared to others, but the fed-batch operation of 40 + 20 + 20 has the higher astaxanthin content of 3.44 mg/g obtained. Overall, the fed-batch operation of 40 + 20 + 20 can lead to the highest total astaxanthin production of 51.22 mg/L.
Table 2
Comprehensive data of batch and fed-batch operation at final glucose concentration of 80 g/L in the 5L fermenter
Glucose concentration (g/L) | Max. biomass (g/L) | Astaxanthin content (mg/g) | Astaxanthin Yield (mg/L) | Astaxanthin Productivity (mg/L/hr) |
|---|
80 | 16.10 | 2.55 | 41.06 | 0.570 |
60 + 20* | 14.01 | 2.89 | 40.49 | 0.562 |
40 + 20 + 20** | 14.89 | 3.44 | 51.22 | 0.711 |
| *feeding at 24th hr |
| ** feeding at 12th and 24th hr |
It seems a low and stable glucose concentration will be beneficial to astaxanthin production, which is often observed in the recombinant microbial cell with rearranged metabolic network conferring improved biosynthetic capability of various bio-products. A key challenge on how to develop an efficient strategy to maximize target product synthesis without reducing cell growth is still a debate topic, since simple knockout or overexpression of essential genes can easily impair the intracellular metabolic network, resulting in metabolic disturbance, accumulation of intermediates and byproducts [11]. To address these problems, a so-called glucose repression strategy was adopted in the fed-batch operation to maximise the cell growth and the recombinant products production. Conclusively, the best way to perform the fed-batch operation for astaxanthin production is using the on-line glucose concentration monitoring system to maintain the designed glucose concentration by continuously feeding the glucose solution. Unfortunately, such a kind of glucose monitoring system is still not easy to obtain in the industry.
Comparison of Astaxanthin production in 5L and in 30L bioreactors
For the purpose of commercialized production of astaxanthin, the scale up fermentation of recombinant K. marxianus 929 Asta strain D56 was performed in a 30L in-situ fermenter. The major difference between in-situ 30L fermenter and 5L desk-top fermenter is the material of the reactor. It is well known that the high DO-level will lead to a high-cell density cultivation. The 5L fermenter is made of glass, which is not able to use tank pressure to increase dissolved oxygen. However, the in-situ 30L fermenter is a stainless tank that can tolerate the high tank pressure to reduce the agitation speed. To further confirm this recombinant K. marxianus can be a candidate for astaxanthin production in the industry. The batch fermentation of K. marxianus at 0 and 1 atm are examined in a 30L in-site fermenter as compared to that of bath operation in a 5L fermenter. The results of 30L operation were shown in Fig. 6, and the comparison data was shown in Table 3. Surprisingly, the biomass in the 5L and in the 30L fermenter didn’t yield a significant difference. However, the astaxanthin content in the 30L operation will be significantly higher than that of 5L operation. A high DO-level in the 30L fermenter is relatively easy to be maintained at the high level as compared to the 5L desk-top fermenter. It is suggested that a high level of DO might be beneficial to the formation of recombinant astaxanthin production. However, due to the limitation of glucose obtained in the batch operation, the biomass didn’t yield any differences among all batch operations in the 5L and in the 30L bioreactor. Conclusively, a high DO condition will be beneficial to the enhancement of astaxanthin content. However, if both the high biomass and the high astaxanthin content were being achieved, a fed-batch operation by maintaining glucose level during the fermentation is suggested.
Table 3
Effects of tank pressure at 5L and 30L agitation fermenter on biomass and astaxanthin production
Fermentation type | Biomass (g/L) | Astaxanthin (mg/g) |
|---|
5L fermenter at 0 atm | 16.10 ± 0.75 | 2.55 |
30L fermenter at 0 atm | 14.63 ± 0.88 | 4.03 |
30L fermenter at 1 atm | 15.96 ± 0.36 | 4.38 |
Table 4
Comparison table of biomass, astaxanthin content and productivity in literature
Microorganism | Bioreactor type | Max biomass (g/L) | Astaxanthin content (mg/g) | Astaxanthin productivity (mg/L/d) | Reference |
|---|
C. zofingiensis ATCC 30412 | 1 L agitation | 12.9 | 1.1 | 1.7 | [15] |
C. zofingiensis ATCC 30412 | 3.7 L agitation | 51.8 | 0.63 | 5.4 | [16] |
C. zofingiensis CCAP 211/14 | Bubble column | 7 | 6 | 1.3 | [17] |
C. glutamicum ASTA | 2 L agitation | 57 | 1.8 | 36 | [18] |
H. pluvialis NIES-144 | 1 L airlift | 3.0 | 14 | 10.5 | [19] |
H. pluvialis M1 | 100 L flat | 3.11 | 55.12 | 8.6 | [20] |
H. pluvialis SAG 34 − 7 | 1 L bubble-flat | 1.59 | 32.1 | 9.7 | [21] |
K. marxianus Sm23 | 5 L agitation | 11.42 | 9.97 | 22.77 | [14] |
X. dendrorhous ZJUT46 | 2000L agitation | 17.42 | 2.27 | 7.2 | [22] |
K. marxianus 929 Asta D56 | 5 L agitation | 16.10 | 2.55 | 13.68 | this study |
K. marxianus 929 Asta D56 | 30 L agitation | 15.96 | 4.38 | 23.30 | this study |
{kind=link}