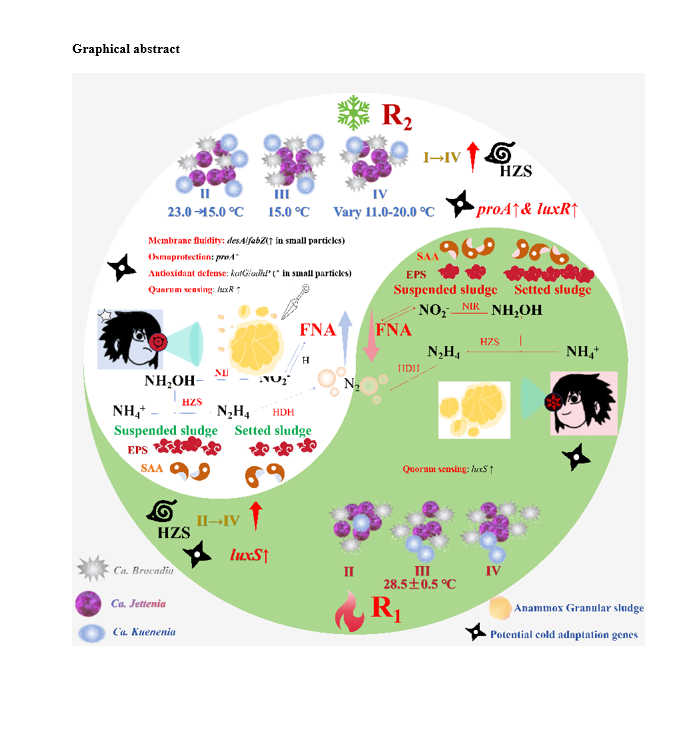
The adaptation mechanisms of anammox granular sludge under fluctuating low-temperature conditions require further elucidation, particularly concerning the interplay between sludge size, microbial community dynamics, and functional gene including nitrogen-removal and cold resistence genes. This study investigated two anammox reactors operated at high-temperature (28.3°C) and fluctuating low-temperature (11–23°C) conditions to elucidate these mechanisms. Results demonstrated that FNA (> 0.5 µg L− 1), rather than free ammonia (FA), was the primary inhibitor of anammox activity at low temperatures, reducing nitrogen removal efficiency by 16% at 15 ℃. Apparent temperature coefficient (Q10) ranged from 0.90 to 1.27, indicating typical temperature-dependent bacterial activity. Low temperatures increased EPS structural vulnerability, weakened granular strength and increased small particles (< 0.25 mm) by 32% while decreasing biomass content (VSS/SS to 27.9%). Our findings demonstrated that the system does not merely undergo inhibition but orchestrates a sophisticated adaptive response: larger granules (> 2 mm) serve as stable reservoirs for core anammox bacteria (notably Candidatus Jettenia caeni), ensuring biomass retention, while smaller granules (< 0.25 mm), generated through low-temperature-induced fragmentation, function as specialized pioneers. These small granules uniquely enrich a suite of cold-adaptation genes, empowering them to directly combat cold stress by modulating membrane fluidity, synthesizing compatible solutes, enhancing antioxidant defense, and fine-tuning cell-to-cell communication, suggesting heightened environmental sensitivity.
Research Article
Size-dependent performance and adaptive mechanisms of anammox granular sludge under fluctuating low-temperature conditions
https://doi.org/10.21203/rs.3.rs-8311852/v1
This work is licensed under a CC BY 4.0 License
You are reading this latest preprint version
Anammox
Low-temperature adaptation
Granular sludge
Bacterial community
Nitrogen removal
Cold resistence genes
Anaerobic ammonia oxidation (anammox) is a highly promising biological nitrogen removal technology for treating wastewater characterized by low C/N ratios and high NH4+-N concentrations. Through the metabolism of anammox bacteria (AnAOB), NO2−-N and NH4+-N are converted into N2 gas under anaerobic conditions (Huo et al. 2020; Liu et al. 2021; Chang et al. 2025). Unlike traditional biological nitrogen removal processes, anammox system requires no organic carbon input and produces lower residual sludge (An et al. 2013; Luyara et al. 2018; Wang et al. 2018; Chang et al. 2025). To date, anammox has been extensively studied and applied for low C/N wastewater treatment (Wang et al. 2024).
Temperature is a critical role in the nitrogen removal performance of the anammox process, and the optimal operation temperature ranged from 30 to 40°C (Jin et al. 2012; Huo et al. 2020; Talan et al. 2021, Zhang et al. 2025). Low temperatures represent one of the major challenges, significantly reducing anammox activity, impairing sludge biomass retention, altering microbial community structure and reducing nitrogen removal efficiency (Wang et al. 2024; Xiong et al. 2025; Zhang et al. 2025). When operated at an environmental temperature of 24.9 ± 1.5°C, the anammox system exhibited significantly lower ammonium removal efficiency (ARE), total nitrogen removal rate (TNRR), and total inorganic nitrogen removal efficiency compared to operation at 35°C (Munoz-Palazon et al. 2025).
When the temperature decreased from 35 to 20°C, the anammox system retained the persistence of AnAOB like Ca. Brocadia and partial denitrification genera (Luyara et al. 2018). When the temperature drops below 15°C, AnAOB activity and ammonia oxidationbacteria (AOB) is severely inhibited (Zhang et al. 2020; Xiong et al. 2025). Liu et al. (2021) observed weakened quorum sensing at 15°C, leading to deteriorated operational performance. AnAOB exhibit limited substrate tolerance at low temperatures (e.g., 12°C) with dominant Ca. Kuenenia (He et al. 2018). Collectively, these studies highlight the significant impacts of low-temperature environments on both microbial community structure and total nitrogen removal efficiency (TNRE) in anammox processes. Xiong et al. (2025) alleviated the oxidative damage of ROS to anammoxosome membrane by ascorbic acid.However, there is still limited knowledge about the low-temperature adaptation mechanisms of anammox bacteria.
The performance of anammox systems is significantly affected by sludge particle size, which affects biomass retention, substrate diffusion, and microbial community structure. Smaller sludge generally had poor settleability and was easily to be washed out of reactor (Zhu et al. 2018; Kang et al. 2023; Chen et al. 2025). The optimal size range of anammox granular sludge (AGS) is operationally dynamic, spanning from micro- to millimeter scale, and is predominantly governed by key operational parameters including substrate characteristics, hydrodynamic shear forces, reactor configuration, and nitrogen loading rates (Zhu et al. 2018; Kang et al. 2023; Pan et al. 2025). To enhance the resistance of AnAOB to low temperatures, gels (composed of polyvinyl alcohol & sodium alginate) were used to immobilize anammox sludge and enhance particle size (Zhang et al. 2025). In one-stage partial nitritation / anammox system, the changes in granular size of granular sludge were reported under 15°C (Chen et al. 2025). However, the evolution of sludge size with the microbial communities, and functional genes including nitrogen-removal and cold resistence genes in different granular size of anammox sludge under fluctuating low-temperature conditions remains poorly understood.
In this study, two anammox reactors operating under distinct temperature regimes were constructed to investigate these aspects. The objectives of this study were to (1) compare the operational performance and stability of anammox systems under high-temperature (28.3°C) and low-temperature (11–23°C) conditions; (2) investigate the characteristics of anammox sludge under temperature variations; and (3) elucidate the evolutionary dynamics of functional bacteria and genes including nitrogen-removal and cold resistence genes in AGS at different temperatures and sludge sizes, revealing the adaptive mechanisms of microbial communities under low-temperature stress.
Synthetic wastewater
Synthetic wastewater was used as the influent for anammox reactors. (NH4)2SO4 and NaNO2 were added as NH4+-N and NO2−-N sources, respectively. Other components included NaH2PO4 (0.01 g L− 1), MgSO4·7H2O (0.59 g L− 1) and CaCl2 (1.00 g L− 1) and EDTA chelated trace elements including Fe(II), Zn(II), Co(II), Mn(II), Cu(II), Mo(VII), Ni(II) and B(III). Levels of trace elements were same to Chang et al. (2025).
Experimental setup and procedures
Two lab-scale UASB reactors, R1 and R2, were constructed with an effective volume of 1.2 L each. The operation of R1 was divided into four operation stages (I, Ⅱ, Ⅲ and Ⅳ). In stage I, reactor R1 was operated at water temperature (23–27°C) for AGS cultivation. The hydraulic retention time (HRT) of UASB was 1.15 h. At the end of stage I (1-84d), the AGS of R1 was evenly mixed, and half sludge was removed into reactor R2. The detail operational parameters of both reactors are presented in Table 1.
······················Table 1········&mi
Water quality analysis
NH4+-N, NO2−-N, NO3−-N and pH of water samples were determined by phenol-hypochlorite spectrophotometry, N-(1-naphthalyl)-ethylenediamine spectrophotometry method and ultraviolet spectrophotometry method, respectively (APHA, 2005). A pH meter (METTLER TOLEDO, Shanghai) was used for pH determination.
Sludge characteristics analysis
EPS extraction and 3D-EEM analysis
AGS samples were obtained at the end of the experiment. The soluble EPS (S-EPS), loosely-bound EPS (LB-EPS) and tightly-bound EPS (LB-EPS) were analyzed based on the method provided by Zhang et al. (2024). Detailed steps are provided in the Supplementary materials. The protein (PN) and polysaccharides (PS) of different EPS were determined by improved Lowry Folin method and Anthrone Method. Three-dimensional excitation-emission matrix (3D-EEM) analysis was performed according to Zhang et al. (2024).
Granular sludge size and settling performance of AGS
At the end of the experiment, AGS of both reactors was collected uniformly and screened by standard sieve. The SS and VSS of AGS with size of < 0.25mm, 0.25-0.5mm, 0.5-1mm, 1–2 mm and > 2 mm were obtained by gravimetric method. The sludge volume after 5minutes (SV5) and 30 minutes (SV30) and SVI of AGS in both reactors were determined according to APHA (2005).
SAA determination
To minimize sludge consumption and its potential impacts on anammox system performance, AGS samples were only obtained for SAA analysis at the end of experiment. SAA values of sludge with different sizes were calculated by matrix concentration curves (Zhang et al. 2024).
Metagenomic sequencing
Metagenomic sequencing was conducted for analyzing the microbial community structure and function genes in AGS Day 7 of Stage II in R1 and R2 (R1-I and R2-I) and at end of stage Ⅱ (R1-Ⅱ and R2-Ⅱ), Ⅲ (R1-Ⅲ and R2-Ⅲ) and Ⅳ (R1-Ⅳ and R2-Ⅳ), and AGS with sizes of < 0.25 mm, 0.25–0.5 mm, 0.5-1mm, 1–2 mm, and > 2 mm at the end of stage Ⅳ in both reactors. The extraction of total community genomic DNA was conducted utilizing the E.Z.N.A. Soil DNA Kit (Omega, M5635-02, USA). Metagenomic sequencing analysis was conducted by Sangon Biotech (Shanghai) Co., Ltd. Species annotation information and functional annotation information of genes were obtained by comparing gene set with NR, ARDB, CAZy, eggNOG, KEGG and SEED databases using DIAMOND (v0.8.20) (Chang et al. 2025).
Calculations
Apparent temperature coefficient (Q10) calculation
The general model for the calculation of Q10 is expressed by the following Eq. 1:

where T is the temperature (°C), R1 and R2 are the TN removal rate (TNRR) (kg m− 3 d− 1) at temperature T1 and T2, respectively.
Free ammonia (FA) and free nitrous acid (FNA) calculation
FA andFNA levels play important roles in the operating stability of anammox system. FA and FNA levels were calculated by the formulas provided in the literature (Anthonisen et al. 1976).
Statistical Analysis
Continuous variable data are presented as mean ± standard deviation. Student's t-test was used to compare the differences between two groups, with a p < 0.05 considered statistically significant. Pearson correlation analysis was applied to assess linear relationships between variables. Quantitative relationships between variables were modeled using simple and multiple linear regression analyses, and the goodness of fit for these models was evaluated using the coefficient of determination (R2). All statistical analysis were performed using OriginPro 2021.
Operation performance of anammox systems under different temperatures
The operational performance of the reactor in stage I is shown in Fig. 1. In the first 58 d (stage I-1), the NH4+-N, NO2−-N and TN removal efficiencies were 96.13% ± 1.30%, 99.36% ± 0.68% and 89.41% ± 1.20%, respectively. At stage I-2, the operation performance of anammox system was not significantly affected, with NH4+-N, NO2−-N and TN removal efficiencies of 93.22%, 99.19%, and 88.81%, respectively. The operation performance of anammox system would not be significantly affected at temperature of 23–26℃ (Liu et al. 2021), but fluctuated nitrogen removal was observed by decreasing temperature from 33℃ to 23℃ in anammox systems (He et al. 2018). Performance recovered to baseline levels after continued operation, suggesting the robustness of the process.
······················ Fig. 1······················
In stages Ⅱ-Ⅳ, R1 had higher average NH4+-N, NO2−-N and TN removal efficiencies of 95.06%±0.94%, 98.38%±1.57% and 86.84%±0.82%, respectively. Operating at low temperatures, R2 experienced a decline in performance, particularly when temperature dropped to 15°C. The average TNRE in R2 decreased to 70.08% ± 8.17% at 11–15°C, highlighting the inhibition effects of low temperature. In stage Ⅲ (135-152d), R2 operated at 15 ℃ exhibited markedly reduced efficiency relative to stage Ⅱ. The average NH4+-N, NO2−-N and TN removal efficiencies were 68.41%±7.97%, 69.95%±9.27% and 61.13%±7.76%, respectively. Due to robust nitrogen removal in stage Ⅳ, R1 exhibited stable operation performance.
In stage Ⅳ, the influent NH4+-N and NO2−-N of R2 reduced to 70 mg L− 1 with average NLR and NRR values of 7.30 ± 0.15 and 5.09 ± 0.54 kg m− 3 d− 1, respectively. Low temperature inhibited anammox activity and deteriorated nitrogen removal (Zekker et al. 2020), and TN removal efficiency of anammox system at 15℃ decreased to 66% (Liu et al. 2021). In this experiment, the TN removal efficiencies at 15℃ ranged from 66.82% to 78.93%. AnAOB had good adaptability to low temperature, but the anammox activity was significantly inhibited below 15℃ (Liu et al. 2020). In this experiment, the anammox activity of the system was significantly inhibited with the average TNRE of 70.08%±8.17% at 11–15℃.
Sludge characteristic analysis
Sludge sizes distribution
In R1 and R2, the dominant sludge fraction consisted of AGS larger than 2 mm, representing 54.65% and 52.49% of total sludge mass and VSS/SS ratios of 77.90% and 78.54%, respectively (Table 2). This similarity suggests that temperature had negligible influence on the VSS fraction of large granules (> 2 mm) (p > 0.05), indicating that the organic matter content of large particles is less affected by fluctuating low temperature of 11–20 ℃ in stage IV. In one-stage partial nitritation/anammox system, elevated aeration intensity induced granular size reduction, with subsequent temperature decrease leading to stabilization at 386 µm (Chen et al. 2025). This study maintained an anammox operation without aeration, consequently preventing the reduction of larger sludge particles even operated at low temperature in R2.
······················Table 2······················
In contrast, the proportion of smaller granular sludge (< 0.25 mm) differed between reactors, accounting for 15.02% in R1 and 19.85% in R2. These finer particles exhibited significantly higher organic content in R1 (VSS/SS = 41.64%) compared to R2 (VSS/SS = 27.94%) (p < 0.01), demonstrating temperature-dependent variation. This trend persisted across granular sludge with other size fractions, and R1 consistently showed higher VSS/SS ratios than R2. The observed reduction in VSS/SS at lower temperatures aligns with reported findings by He et al. (2018). Chen et al. (2025) reported that larger granular sludge contained more abundant and sizable inorganic cores, ensuring superior size maintenance and settleability compared to smaller granules with fewer and smaller cores. In this study, low temperature reduces the proportion of organic matter in small particle sludge, suggesting reduced bacterial or EPS composition of small particle sludge.
The VSS/SS ratio (%) exhibits a linear correlation with particle size (median size of AGS; d, mm) have a linear correlation (Fig. S1). The VSS/SS reduction rates in reactor R1 and R2 were 17.82 mm− 1 and 21.26 mm− 1, respectively. Compared to the higher temperature in R1 (28.3 ± 0.5 ℃), the fluctuating low temperature (10–20 ℃, stage IV) significantly decreased the VSS/SS reduction rates in granular sludge with size < 2 mm. These results showed that low temperatures not only diminished biomass content in AGS but also promoted the formation of smaller particles (< 0.25 mm).
Sludge settling performance analysis
Granular sludge has the advantages of good compactness, excellent settling ability, strong resistance to impact loads and high microbial activity (An et al. 2013). However, a portion of AGS tends to become buoyant due to decreased density after long-term operation, which will affect the retention capacity of sludge, and consequently deteriorates operational performance (Chen et al. 2014; Tang et al. 2017; Tao et al. 2020). Low-temperature operation led to the increased production of suspended AGS. After 30 minutes of settling, the volume ratios shifted to 7% (suspended AGS) and 25% (settled AGS) for R1, and 12% (suspended AGS) and 33% (settled AGS) for R2.
SAA analysis
SAA of AGS with different granular sizes As shown in Fig. 2(a), all AGS in R1, regardless of sludge size, showed good anammox activity. Especially, the sludge with sizes of 0.25-0.5mm and < 0.25mm had higher activity with SAAs of 68.49 and 66.4 mg TN g-1 VSS h-1. Generally, smaller AGS exhibited higher biological activity due to their larger specific surface area and reduced mass transfer resistance (An et al. 2013). Consistent with this observation, the sludge < 0.5 mm in this experiment displayed the highest anammox activity. In R2, AGS with granular size of > 2mm, 0.5-1mm and < 0.25mm showed similar activity to R1. The SAA value of AGS with granular size of 1-2mm was 3.88 mg TN g-1 VSS h-1, which was significantly less than 8.91 mg TN g-1 VSS h-1 of R1. However, the SAA value of AGS with size of 0.25-0.5mm in R2 was only 1.42 mg TN g-1 VSS h-1, which was nearly negligible compared with 68.49 mg TN g-1 VSS h-1 of AGS in R1 with same granular sizes. This phenomenon may be caused by the fact that the 0.25–0.5 mm granules, likely resulting from the fragmentation of larger, more protected granules, were subjected to a severe dual stressor of high concentrations of inhibitory FNA (Section 3.2) and a reduction in biomass quality (lowest VSS/SS ratio of 25.0%, Table 2). This stark contrast underscores a size-dependent ecological stratification within the granular sludge community under low-temperature stress, where the 0.25–0.5 mm fraction represents a vulnerable, transitional state susceptible to disintegration and inhibition, while the < 0.25 mm fraction embodies a resilient, highly specialized pioneer community.
······················ Fig. 2······················
SAA of suspended sludge and settled sludge The SAA values of suspended sludge and settled sludge in R1 and R2 are shown in Fig. 2(b). The SAA values of suspended sludge and settled sludge in R1 were 18.05 and 17.75 mg TN g-1 VSS h-1, respectively. Corresponding values in R2 were 7.47 and 9.77 mg TN g-1 VSS h-1, respectively. Experimental results showed that suspended sludge and settled sludge in R1 had higher anammox activity.
EPS analysis
EPS contribute to cell adhesion and anaerobic granulation (Tang et al. 2017; Wang et al. 2020a). In this study, negligible amounts of PN and PS were detected in S-EPS and LB-EPS. In TB-EPS (Fig. 3), higher PN levels were associated with AGS having granular sizes of > 0.5 mm. The maximum PN of 894.5 mg g− 1 VSS was obtained in AGS with size of 1-2mm sludge in R1, and 562.1 mg g− 1 VSS in AGS with size of > 2mm in R2. In contrast, the maximum PS content was found in 1–2 mm AGS, with values of 109.2 mg g− 1 VSS (R1) and 98.6 mg g− 1 VSS (R2).
······················ Fig. 3······················
PN/PS ratios in AGS with sizes > 2mm, 1–2 mm, 0.5-1 mm, 0.25–0.5 mm and < 0.025 mm for R1 were 8.5, 8.2, 7.2, 11.0, and 4.0, respectively. Corresponding values for R2 were 6.0, 5.6, 6.8, 7.0, and 3.6, respectively. These values align well within the typical range of 3.4–12.1 reported for EPS in previous studies (Lotti et al. 2019, Feng et al. 2019; Wang et al. 2020a; Zhang et al. 2024). PN/PS ratio is commonly used to characterize the strength and settling performance of AGS, and a lower PN/PS ratio indicates a better settling performance and higher strength of AGS (Wang et al. 2020a,b). In this study, low operational temperatures significantly reduced these ratios, indicating EPS structural vulnerability. TB-EPS and PN appear particularly crucial for maintaining AGS structure (Wang et al. 2020a,b), and hydrophobic amino acids and loose configuration of PN facilitated anammox sludge aggregation (Hou et al. 2015). Higher PN/PS values in R1 suggested a better operation performance and settlement capability of AGS.
Suspended sludge had a higher EPS content compared to AGS with good settling performance (Chen et al. 2014). As shown in Fig. 3, suspended and settled sludge in R1 was relatively compact, with no detectable S-EPS or LB-EPS under the experimental conditions. In contrast, the sludge in R2 was relatively loose due to low temperature, and both S-EPS and LB-EPS of the settled sludge contained detectable levels of PN (18.47 and 41.58 mg g⁻¹ VSS, respectively) and PS (13.81 and 38.84 mg g− 1 VSS, respectively). Higher PN/PS ratios suggested lower strength of granular sludge (Wang et al. 2020a). The significantly higher PN/PS ratio in reactor R1 compared to R2 (p < 0.05) (Fig. 3) suggested that low-temperature conditions induced structural fragility in granular sludge. EPS was used by cold adapted microorganisms to maintain stability and protect cellular structures (Guo et al. 2025). PS in EPS can serve as natural cryoprotectants for cells to adapt to low temperatures. This protective effect in this study may be achieved by PS in TB-EPS. Meanwhile, the PS value in TB-EPS of settled sludge in R2 (183.4 mg g− 1 VSS) was higher than R1 (132.9 mg g− 1 VSS). PS content was identified as a primary factor influencing anammox sludge granulation, even PS levels much less than PN (Zhu et al. 2018). The higher PS levels observed in R2 in this study indicated that the reactor maintained good granulation.
The 3D-EEM of EPS in R1 and R2 reactors revealed distinct compositional patterns between sludge fractions (Fig. S2-S3). While no characteristic peaks were detected in S-EPS or LB-EPS, and TB-EPS exhibited multiple fluorescence peaks across all samples (S2 section in Supplementary materials). Notably, granular sludge > 0.5 mm showed significantly higher PN content than smaller fractions, suggesting particle size-dependent EPS composition.
FA and FNA analysis
The FA concentration of influent and effluent of R1 and R2 are shown in Fig. 1 (d). Studies have shown that FA and FNA easily cross the cell membrane and dissolve in lipids, and NH4+-N and NO2−-N inhibit AnAOB through FA and FNA (Jin et al. 2012; Talan et al. 2021). FA does not inhibit AnAOB at concentrations < 2 mg L− 1 (Jung et al. 2007), and the inhibition thresholds of FA were 13 mg L− 1 (Waki et al. 2007) or 20 mg L− 1 (Fernández et al. 2012). In this experiment, effluent FA values of R1 and R2 were 0.70 ± 0.42 and 1.30 ± 0.65 mg L− 1, respectively. Except for some days of R2, the effluent FA levels of both reactors were below threshold value of 2 mg L− 1, indicating that neither reactor may be inhibited by FA.
When FNA was lower than 0.5 µg L− 1, anammox can maintain good performance (Jin et al. 2012). In order to maintain the stable operation of anammox process, FNA should not be higher than 0.5µg L− 1 (Fernández et al. 2012). In this experiment, the effluent FNA values of reactor R1 were maitained at lower levels in stages Ⅱ (0.15 ± 0.26 µg L− 1), Ⅲ (0.54 ± 0.27 µg L− 1) and Ⅳ (0.18 ± 0.13 µg L− 1) (Fig. 1(e)). The effluent FNA of R2 maintained higher levels in stages Ⅱ (0.69 ± 0.51 µg L− 1) and Ⅲ (1.78 ± 0.59 µg L− 1). Even influent NH4+-N and NO2−-N decreased from 104.3 ± 1.98 and 100.0 ± 1.03 mg L− 1 (Stage Ⅲ) to 74.4 ± 2.78 and 71.2 ± 1.73 mg L− 1 (Stage Ⅳ) (Table 1), the FNA level remained high at 0.63 ± 0.39 µg L− 1 (Stage Ⅳ). These vlues were higher than the reported FNA inhibition threshold of 0.5µg L− 1 (Fernández et al. 2012), indicating a potential inhibited by FNA.
······················Table 2······················
Apparent temperature coefficient analysis
According Eq. (1), pseudo-steady-state data of reactor R2 were used with fitting curves of ln(R2/R1) = 0.1161 (T2-T1)/10–0.0056 (R2 = 0.6026) (R1, I-2), ln(R2/R1)=-0.1078(T2-T1)/10 + 0.0386 (R2 = 0.7342) (R2, II), and ln(R2/R1) = 0.2373(T2-T1)/10 + 0.0211 (R2 = 0.6030) (Ⅳ). The slope values can be used for Q10 calculation, and apparent temperature coefficient Q10 were calculated to be 1.12, 0.90 and 1.27 at phases I-2 (R1), II (R2) and Ⅳ (R2), respectively. The temperature coefficient Q10 is usually greater than 1. However, in this study, the Q10 of reactor R2 at phase II was 0.9. Wang et al. (2024) reported the increased anammox activity from 39.7 to 45.5 mg N g− 1 VSS d− 1 when temperature decreased from 30 to 15 ℃ due to the endogenous denitrification, higher AnAOB retention in biofilm and a continuous nitrite supply. Obviously, the decrease in anammox activity in this study did not caused by the increase in temperature, but may be due to the inhibitory effect of FNA (Section 3.3).
Bacterial community analysis
Bacterial community composition in AGS across different size fractions
Alpha diversity analysis revealed distinct size-dependent patterns in granular sludge. Smaller AGS exhibited a significantly higher gene number compared to larger granules (Table S1). As shown in Fig. 4, Proteobacteria had highest RA in small AGS, with RAs of 31.66% (< 0.25 mm) and 29.35% (0.25–0.5 mm) in R1, compared to R2 with higher values of 34.39% and 32.24%, respectively. Planctomycetes predominated in larger granules, with RAs of 39.15% (0.5-1 mm), 29.91% (1–2 mm) and 38.82% (> 2 mm) in R1, while R2 exhibited values of 27.32%, 34.74% and 39.17% for corresponding size fractions. Both Proteobacteria and Planctomycetes contain numerous nitrogen-removing bacteria. In addition, Chloroflexi is an important accompanying functional bacterial community and serves dual functional roles in sludge systems: (1) providing structural support as a skeletal framework, and (2) decomposing and utilizing macromolecules released from lysed cells (He et al. 2018; Liu et al. 2021). Meanwhile, Chlorobi significantly contributes to extracellular protein degradation processes (Zhao et al. 2018).
······················ Fig. 4······················
As shown in Fig. 4, the small sludge fraction (< 0.25 mm) exhibited the highest genus diversity. Notably, three AnAOB genera, including Ca. Jettenia, Ca. Brocadia and Ca. Kuenenia, consistently dominated the bacterial communities in both reactors. Except for the 1–2 mm fraction, the RAs of Ca. Jettenia in R1 were significantly higher than R2 (p < 0.05). This result showed that Ca. Jettenia is more sensitive to temperature changes, low temperature decreased RA in AGS. Notably, Ca. Brocadia was dominant in the 0.5-1 mm fraction in R1 and in the > 0.5 mm fractions in R2. The RAs of Ca. Brocadia in R1 were significantly higher than those in R2 for granular sizes ≤ 2 mm (p < 0.05) but significantly lower in > 2 mm AGS (p < 0.05). This result showed that Ca. Brocadia thrives in mid-sized granules at ambient temperatures (R1) but shifts to larger granules in cold conditions (R2). There is no significant difference of Ca. Kuenenia between reactors R1 and R2 across the granular sizes (t-test, p > 0.05), suggesting Ca. Kuenenia appears more versatile, with minor temperature-dependent shifts.
Other bacterial genera related to nitrogen removal were also found in both reactors, including Nitrosomonas (AOB), Nitrobacter (NOB) and denitrifying genera such as Thauera, Dechloromonas, Bacillus, Sporosarcina, Clostridium and Gordonibacter, etc. Tao et al. (2020) reported that sludge with large granular sizes favored the enrichment of denitrifying bacteria. Under constant higher temperature conditions in R1, the total RA of denitrifying genera in AGS varied across different size fractions, and the highest RA of denitrifying genera was observed in the 1–2 mm sludge fraction, suggesting a preference for larger granules. However, there was no significant difference in RA of denitrifying genera in different sludge sizes of R2.
Bacterial community structure in AGS across different operational stages
In R1, all diversity indices remained statistically consistent (p > 0.05), and temperature fluctuations and low temperature significantly affected diversity (p < 0.05) (Table S1). Planctomycetes, Proteobacteria and Chloroflexi were the dominant phyla in AGS across all four stages (Fig. 4). In R1, the RAs of Planctomycetes progressively decreased from 34.68% (initial days of stage II) to 20.28% (stage IV). In contrast, R2 maintained consistently higher Planctomycetes RAs of 38.56%, 38.09%, 39.35% and 32.91% in stages I-IV, respectively. He et al. (2018) reported that Planctomycetes exceeded Proteobacteria by 56.34% at 13°C. Throughout all stages in R2, Planctomycetes remained dominant, suggesting it may possess a competitive advantage at low temperatures. Chloroflexi abundances in R1 (5.91%-6.97%) were generally higher than in R2 (6.14%-6.47%) during stages I-III, but lower in stage IV (5.91% vs 6.43%). Liu et al. (2021) reported that RA of Chloroflexi was 12.36% at 25℃ and 8.47% at 15℃. In this study, the decreased temperature did not decrease the RA of Chloroflexi in R2.
There were three dominant anammox genera including Ca. Jettenia, Ca. Brocadia and Ca. Kuenenia. Notably, R1 displayed distinct bacterial community structures across different phases. Previous studies have demonstrated that Ca. Brocadia dominates in temperature ranges of 6–25℃, indicating its psychrotolerant characteristics (Huo et al. 2020; Liu et al. 2020; Wang et al. 2024). Additionally, research suggests that lower temperatures may enhance the accumulation of Ca. Kuenenia (He et al. 2018; Liu et al. 2021). Ca. Kuenenia and Ca. Brocadia have been reported to play a critical role in maintaining higher nitrogen removal efficiency in anammox systems under low-temperature conditions (Zhang et al. 2025). However, in our experimental system operating at 11–23℃ (R2), both Ca. Jettenia and Ca. Brocadia emerged as dominant genera in all operational phases. The RA of Ca. Jettenia ranged from 16.08% to 23.73%, consistently exceeding that of Ca. Brocadia (8.25%-15.53%). These findings contrast revealed that Ca. Brocadia retains its low-temperature adaptability, while Ca. Jettenia also exhibits notable psychrotolerant capabilities under our experimental conditions.
Dynamic response and particle size distribution characteristics of AnAOB species under low temperature conditions
As shown in Fig. 4e and Table S2, the dominant AnAOB species in different AGS samples were Ca. Kuenenia stuttgartiensis, Ca. Brocadia sinica, Ca. Brocadia fulgida and Ca. Jettenia caeni. Ca. Jettenia caeni and Ca. Kuenenia stuttgartiensis were reported as dominant anammox species treating low-level NH4+-N wastewater (Ma et al. 2017). In contrast, when treating high-substrate wastewater, Ca. Brocadia were dominated with Ca. Scalindua rarely detected (Zhu et al. 2018). This study revealed significant differences compared to previously reported findings. Experimental results revealed significant distribution differences among AnAOB species across operational stages and AGS sizes. Ca. Brocadia fulgida in R1 showed a obviouosly increase in RA from 5.74% (initial day of stage II) to 9.36% (stage IV) (p < 0.05), and Ca. Brocadia sinica exhibited a similar increasing trend. Due to lower operation temperature, Ca. Jettenia caeni and Ca. Kuenenia stuttgartiensis in R2 decreased by 14.1% and 39.8%, respectively, from initial days of stage II to stage IV.
Granular size analysis demonstrated distinct size selectivity in distribution of AnAOB species. Pearson's correlation analysis (using median of granular size of each fraction as an independent variable), Ca. Jettenia caeni exhibited a strong positive correlation with granular size in both reactors R1 (r = 0.92, p < 0.01) and R2 (r = 0.95, p < 0.01). Its RA reached 23.70% (R1) and 18.07% (R2) in large granular sludge (> 2 mm), significantly higher than the 1.79% and 1.01% observed in small AGS (< 0.25 mm). The dominance of Ca. Jettenia caeni in larger granules (> 2 mm) may be attributed to its ability to create low-oxygen microenvironments within granular cores. Obviously, Ca. Jettenia caeni tends to be accumulated in larger AGS. The maximum value of Ca. Brocadia fulgida in both reactors occurs in AGS with size of 0.5-1 mm (R1:15.89%; R2: 9.16%), and too large or too small size of AGS may be not conducive to its enrichment. This result may suggest a potential upper limit to the size adaptation of Ca. Brocadia fulgida. It is possible to have optimal mass transfer performance at this particle size. However, Ca. Brocadia fulgida in R2 also displayed size dependency (r = 0.87, p < 0.05). Obviously, large AGS may better reduce the impact of low temperature stress on anammox to some extent. Due to operational differences, results of this study deviate from those reported by Zhu et al. (2018), who identified optimal granular sizes of 0.5–0.9 mm in an anammox system dominated by multiple species of Jettenia, Brocadia, and Anammoxoglobus.
Multivariate regression analysis indicated that the RA of Ca. Jettenia caeni was significantly and positively influenced by both particle size and temperature (RA = 0.78D50 + 0.65T − 5.32 (R2 = 0.86, p < 0.01)) (D50: median size of AGS, mm; T, water temperature in reactor, °C). Larger AGS with higher temperatures (> 25°C) promote the enrichment of Ca. Jettenia caeni, whereas low temperatures compromise its competitive advantage. In contrast, the distribution of Ca. Kuenenia stuttgartiensis showed no significant correlation with granular size, but its RA in R2 decreased most sharply (39.8%) under low-temperature conditions, highlighting its enhanced sensitivity to temperature fluctuations. The distribution of AnAOB is co-regulated by operational conditions and granular size. Temperature is a critical determinant of species competitiveness, while particle size further filters community structure by influencing substrate transfer or microenvironments. Optimizing the temperature and the distribution of granular size can enhance the stability of anammox process (Chen et al. 2021; Kang et al. 2023; Zhang et al. 2025).
Functional gene analysis
Functional genes related to nitrogen removal across different operational stages
The nitrite reductase gene (nir), hydrazine dehydrogenase gene (hdh), hydroxylamine oxidase gene (hox) and hydrazine synthase gene (hzs) are all closely related to anammox reaction (Chang et al. 2025). As shown in Fig. 5 and Table S3-S4, the RA of anammox-related genes in R1, including NIR and hdh, remained stable across all stages. However, the RA of hzs increased from 1.93% in initial days of stage II to 4.20% in stage IV. R2 operated at variable low-temperature conditions. When temperature decreased from 23 ℃ to 15 ℃ (stage II), the RA of hzs and hdh decreased, and the RA of NIR increases from 3.44% to 5.05%. In stable temperature (Stage III) and temperature fluctuations (stage IV) resulted in no significant changes in RA of these functional genes. Notably, neither reactor exhibited significant differences in the proportions of nitrification-related functional genes (amoA/B/C, hao, and nxrA/B) across different stages. However, the RAs of denitrification genes (nasA and narG) in R2 were higher than those in R1, suggesting that lower temperatures may promote denitrification activity. Potential endogenous denitrification may be one of the contributing factors to the observed decrease in the VSS/SS ratio (Table 2).
······················ Fig. 5······················
Functional genes related to nitrogen removal across different size fractions
In anammox reaction, the highest RAs of NIR gene were 4.49% (R1) and 3.89% (R2) in sludge with a granular size > 2mm (Fig. 5). The NIR genes, including haem-containing NIR (nirS), copper-containing NIR (nirK) and assimilatory NIR (nirB) genes were consistently higher in R1 compaired with R2. The RAs of hdh were similar with each other in all sludge in R1 and R2. The RA of hzs in sludge of > 2mm was higher in R2 (4.46%) than in R1 (2.67%). However, the RAs of hzs in R1 with other size fractions were higher than R2. In anammox systems, HOX functions as a metabolic decomposition enzyme with a role similar to HAO, recovering hydroxylamine leaked from HZS (Marcel et al. 2018). The RA of hox exhibited remarkable consistency across granular sludge sizes in both reactors, suggesting that hox was uniformly distributed in AGS of all size fractions and remained stable under varying temperature conditions.
The ammonia monooxygenase (AMO) genes were detected at relatively low levels and predominantly existed in smaller AGS. The distribution of HAO gene and NXR gene was similar in both reactors. Notably, the highest RA of HAO gene was observed in sludge with size > 2 mm, with R1 showing levels of 5.61% and 10.79%, while R2 demonstrated comparable values of 5.43% and 9.93%, respectively. The denitrification processes in both reactors involved two distinct nitrate reductase systems: assimilatory nitrate reductases (nirA and nasA) and membrane-bound nitrate reductases (narG). Genes including nasA and narG exhibited comparable distribution patterns across AGS in both reactors, with their highest RAs consistently observed in the 0.5-1 mm sludge fraction, reaching 7.71% in R1 and 6.91% in R2, respectively. Additionally, dissimilatory periplasmic cytochrome c nitrite reductase (nrfA/H) (Marcel. et al. 2018) were also identified in both systems. The similar abundance profiles of nrfA/H between R1 and R2 suggested that these genes remained stable regardless of temperature variations.
Potential cold adaptation genes in AGS
Genes related to regulation of microbial membrane adaptability Increasing unsaturated fatty acids while reducing hydrocarbon chain length serves to preserve membrane fluidity and maintaining cell structure under low temperature conditions (Diomandé et al. 2016; Guo et al. 2025). The RA of cold adaptation genes related to regulation of microbial membrane adaptability indluding fatty acid desaturase genes (desK, desR, desA and desA1) and fatty acid synthesis genes (fabA, fabB, fabM and fabZ) are shown in Fig. 5c and Fig. 5d, respectively. Student t-test showed that there were no significantly different at different operation stages in both reactors (p > 0.05). All genes except fabM (Not detected in large particles) showed significant differences between small (< 0.5 mm) and large particles (> 1mm) (p < 0.05), with small particles exhibiting higher RA values. Statistical analysis confirms that particle size is a critical factor in regulation of cold adaptation gene of fatty acid desaturase genes and fatty acid synthesis genes, while operational stages (temperature differences) had negligible effects.
Genes related to compatible solute genes Compatible solutes, such as trehalose, betaine and proline, are accumulated within cells to inhibit crystallization and counteract the impact of intracellular osmotic pressure (Zhang et al. 2022a; Guo et al. 2025). The RA of compatible solute genes including trehalose biosynthesis genes (otsA and otsB), glycine betaine synthesis genes (betA and betB/gbsA) and proline biosynthesis genes (proA, proB and proC) in different anammox sludge samples are shown in Fig. 5e. In most samples, the RA of proA and proB were significantly higher than proC. Especially, the RA of proA gene in R2 was significantly higher than R1 in stages II, III and IV (0.025%, p < 0.05). The findings indicated that low temperatures may upregulated the genes related to compatible solute synthesis as a response to environmental stress. As the particle size increases, the RA of proA and proB showed an upward trend (e.g., RA of proA in R2 with sludge size of > 2mm is 0.026%), indicating that larger granular sludge may possess a stronger capacity for compatible solute synthesis.
Genes related to cold-adapted enzyme genes Cold-adapted enzyme genes (adhP, adhE, lipB, lipA) and ROS scavenging genes (SOD2 (Fe/Mn SOD), SOD1 (Cu/Zn SOD), and katG) are shown in Fig. 5f. Significantly higher RA levels of adhP and katG were observed in small particles (< 0.25 mm) compared to large particles (> 2 mm). The RA levels of adhP were increased by 108.1% and 123.8% in reactors R1 and R2, respectively. At the same time, RA levels of katG were 2.03 and 2.37 times of that in R1 and R2, respectively. This suggests that bacteria in small-particle sludge enhance cold tolerance through reinforced ethanol metabolism (adhP) and antioxidant defense systems (katG-SOD2). Notably, despite R2 reactor operating at lower temperatures, its RAs of cold-adapted enzyme genes and reactive oxygen species (ROS) scavenging genes showed no significant difference from R1 (p > 0.05). This result indicated that AGS with lower particle sizes upregulated cold tolerence gene, rather than environmental temperature. It has been reported that low-temperature-induced ROS may change the anammoxosome membrane properties, consequently afffecting material and energy metabolism of AnAOB (Xiong et al. 2025). In this study, RAs of SOD2 remained consistently higher levels of 0.030%-0.036% and showed nearly no significant difference across different operational stages or granular sludge sizes in both reactors, indicating minimal influence from temperature and granular sludge size.
Genes related to cold shock proteins genes Cold shock proteins including CspA, CspB and the peptidyl-prolyl isomerase PpiD are important in enhancing the low-temperature adaptability of the cold-shock acclimated sludge (Zhang et al. 2025). In this study, nearly no cold shock proteins genes cspA, cspB and PpiD were observed in any AGS samples. This result showed that the no cold shock proteins may be needed for AGS to adapt the fluctuating low temperature of 11–23°C.
Genes related to quorum sensing regulation genes The operation of the anammox process under low temperature is influenced by quorum sensing regulation (Zhang et al. 2022b). As shown in Fig. 5g, the RA of luxS gene in R1 was consistently higher than in R2 with fluctuations low temperature, indicating that stable high temperature of 28.3 ± 0.5 ℃ favored the enrichment of luxS-associated microorganisms. The primary function of luxS is to synthesize autoinducer-2 (AI-2), a universal quorum-sensing signal molecule widely present in both gram-positive and gram-negative bacteria (Gao et al. 2024; Li et al. 2025). Furthermore, the luxS abundances in small-particle sludge (< 0.25 mm) were significantly higher than in other particle size groups, implying that reduced particle size may enhance quorum sensing activity. In contrast, R2 exhibited higher RAs of luxR/vanR gene in stage III and IV, suggesting that cold stress may trigger specific genetic responses. The LuxR receptor protein forms a complex with AHLs, and this LuxR-AHL complex functions as a transcriptional regulator, either inducing or repressing downstream gene expression when the AHL concentration reaches a critical threshold (Yang et al. 2024), and the expression of luxR was enhance at lower temperatures (Zhang et al. 2023). The anomalously low RA value of luxR/vanR in R2-II (0.00002%) may be related to the gradual decrease in temperature. The distribution of the luxS, luxO, and luxR/vanR genes were similar in the same particle size groups between reactors R1 and R2 (Fig. 5g), suggesting particle size may influence these quorum sensing genes independently of temperature conditions.
Adaptation mechanisms of anammox granular sludge to fluctuating low temperature
Under the stress of fluctuating low temperatures, anammox granular sludge achieves resistance and adaptation through the synergistic evolution of its physical structure, microbial community and functional genes, characterized by a remarkable size-mediated division of labor (Fig. 6). Macrostructure remodeling occurs as low temperatures increased the proportion of small particles (< 0.25 mm) which mitigate substrate mass transfer limitations through their larger specific surface area, showing higher bioactivity. Concurrently, larger granules (> 2 mm) serve as stable reservoirs for core anammox bacteria, with Candidatus Jettenia caeni dominating in larger granules (> 2 mm) and showing a positive correlation with both particle size and temperature, while Candidatus Kuenenia stuttgartiensis exhibits greater sensitivity to temperature fluctuations. At the molecular level, small granules serve as the “front line” of cold resistance, enriching a suite of cold-adaptation genes involved in fatty acid desaturation and synthesis, compatible solute biosynthesis, antioxidant defense, and quorum sensing, enabling them to modulate cell membrane fluidity, maintain osmotic balance, scavenge reactive oxygen species, and enhance cell-to-cell communication. Thus, the anammox system adapts through a collaborative strategy where larger granules maintain biomass and core community stability, while smaller granules function as “pioneers”, directly resisting environmental pressure by activating various molecular-level protective mechanisms.
······················ Fig. 6······················
Anammox activity exhibited a typical temperature dependence with Q10 of 0.90-1.27. FNA (>0.5 μg L-1) was identified as the primary inhibitor of anammox activity at low temperatures, reducing TNRE by 16% at 15 °C. Low temperatures increased EPS structural vulnerability and raised small particles (<0.25 mm) by 32% while decreasing biomass content (VSS/SS to 27.9%). Notably, Ca. Jettenia caeni exhibited strong adaptability to low temperatures and dominance in larger granules, linked to enhanced nitrogen removal. Small granules (<0.5 mm) contained higher abundances of cold-adaptation genes (fatty acid desaturases, compatible solute synthesis, antioxidant defenses and quorum sensing regulation), indicating their environmental sensitivity.
Author contributions Guang-feng Yang: Data curation, Methodology, Investigation, Visualization, Writing-Original draft, review & editing, Resources. Qi Luo: Writing-Original draft, Data curation, Conceptualization. Neng-feng Qian: Data curation, Conceptualization. Chang Ben-ze: Data curation, Writing-Original draft, Visualization, Conceptualization. Ya-lei Liu: Data curation, Writing-Original draft, Visualization, Conceptualization. Resources. Dong-zhi Chen: Supervision, Resources. Li-juan Feng: Methodology, Writing-review & editing, Resources.
Funding This research was supported by Zhejiang Provincial Natural Science Foundation of China under Grant No. LY23E080006 and National Natural Science Foundation of China under Grant No. 51808498.
Competing interests The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethical approval This article does not contain any studies with human participants or animals performed by any of the authors.
Data Availability Statement The data that support the findings of this study are available from the corresponding author upon reasonable request.
- An P, Xu XC, Yang FL, Li ZY (2013) Comparison of the characteristics of anammox granules of different sizes. Biotechnol Bioprocess Eng 18:446-454.
- Anthonisen AC, Loehr RC, Prakasam TBS, Srinath EG (1976) Inhibition of nitrification by ammonia and nitrous-acid. J Water Pollut Control Fed 48:835-852.
- APHA (2005) Standard Methods for the Examination of Water and Wastewater, 21st edn. American Public Health Association, Washington, DC.
- Chang BZ, Huang XL, Chen DZ, Jin RC, Yang GF (2025) How biofilm and granular sludge cope with dissolved oxygen exposure in anammox process: Performance, bioaccumulation characteristics and bacterial evolution. J Environ Manage 373:123986.
- Chen C, Jiang Y, Zou X, Guo M, Liu H, Cui M, Zhang TC (2021) Insight into the influence of particle sizes on characteristics and microbial community in the anammox granular sludge. J Water Process Eng 39:101883.
- Chen H, Ma C, Yang GF, Wang HZ, Yu MZ, Jin RC (2014) Floatation of flocculent and granular sludge in a high-loaded anammox reactor. Bioresour Technol 169:409-415.
- Chen Y, Feng G, Tamaishi M, Li YY (2025) Low temperature impact and HAP-enhanced one-stage partial nitritation/anammox process: Long-term stability, high-rate nitrogen removal, and operational strategies at 15 ℃. Water Res X 28:100340.
- Diomandé SE, Doublet B, Vasaï F, Guinebretière MH, Broussolle V, Brillard J (2016) Expression of the genes encoding the CasK/R two-component system and the DesA desaturase during Bacillus cereus cold adaptation. FEMS Microbiol Lett 363:fnw174.
- Feng CJ, Lotti T, Lin Y, Malpei F (2019) Extracellular polymeric substances extraction and recovery from anammox granules: Evaluation of methods and protocol development. Chem Eng J 374:112-122.
- Fernández I, Dosta J, Fajardo C, Campos JL, Mosquera-Corral A, Méndez R (2012) Short- and long-term effects of ammonium and nitrite on the Anammox process. J Environ Manage 95:S170-S174.
- Gao Y, Peng D, Wang X, Lin S (2024) Effects of the quorum sensing related luxS gene and lsr operon on Klebsiella michiganensis resisting copper stress. Environ Res 256:119244.
- Guo Z, Li YG, Wang ZB, Zhou X, Ni SQ (2025) Adaption strategies of extremophiles and the construction of wastewater treatment systems driven by extremophiles. Environ Res 282:121979.
- He SL, Chen Y, Qin M, Mao Z, Yuan L, Niu Q, Tan X (2018) Effects of temperature on anammox performance and community structure. Bioresour Technol 260:186-195.
- Hou X, Liu S, Zhang Z (2015) Role of extracellular polymeric substance in determining the high aggregation ability of anammox sludge. Water Res 75:51-62.
- Huo TR, Zhao YP, Tang X, Zhao HZ, Ni SQ, Gao Q, Liu ST (2020) Metabolic acclimation of anammox consortia to decreased temperature. Environ Int 143:105915.
- Jin RC, Yang GF, Yu JJ, Zheng P (2012) The inhibition of the Anammox process: A review. Chem Eng J 197:67-79.
- Jung JY, Kang SH, Chung YC, Ahn DH (2007) Factors affecting the activity of anammox bacteria during start up in the continuous culture reactor. Water Sci Technol 55:459-468.
- Kang P, Liang Z, Zhang Q, Zheng P, Yu G, Cui L, Liang Y (2023) The optimum particle size of anaerobic ammonia oxidation granular sludge under different substrate concentrations. J Environ Manage 328:116992.
- Li S, Bai Y, Li Z, Ren NQ, Ho SH (2025) Quorum sensing enhances interspecies electron transport in biological wastewater treatment: Insights from molecular mechanisms and enhancement strategies. Chem Eng J 520:165938.
- Liu LJ, Ji M, Wang F, Tian ZK, Yan Z, Wang SY (2021) N-acyl-L-homoserine lactones release and microbial community changes in response to operation temperature in an anammox biofilm reactor. Chemosphere 262:127602.
- Liu T, Lim ZK, Chen H, Hu S, Yuan Z, Guo J (2020) Temperature-tolerated mainstream nitrogen removal by anammox and nitrite/nitrate-dependent anaerobic methane oxidation in a membrane biofilm reactor. Environ Sci Technol 54:3012-3021.
- Lotti T, Carretti E, Berti D, Martina MR, Lubello C, Malpei F (2019) Extraction, recovery and characterization of structural extracellular polymeric substances from anammox granular sludge. J Environ Manage 236:649-656.
- Luyara DAF, Pereira AD, Leal CD, Davenport R, Werner D, Filho CRM, Bressani-Ribeiro T, Augusto DLCC, De Araújo JC (2018) Effect of temperature on microbial diversity and nitrogen removal performance of an anammox reactor treating anaerobically pretreated municipal wastewater. Bioresour Technol 258:208-219.
- Ma B, Qian W, Yuan C, Yuan Z, Peng Y (2017) Achieving mainstream nitrogen removal through coupling anammox with denitratation. Environ Sci Technol 51:8405-8413.
- Marcel MMK, Hannah KM, Boran K (2018) The microbial nitrogen-cycling network. Nat Rev Microbiol 16:263-276.
- Muñoz-Palazón B, Gallardo-Altamirano MJ, Gonzalez-Lopez J, Gonzalez-Martinez A, Osorio F (2025) Performance and microbiome of an anammox biofilter for treating secondary wastewater: Effect of organic matter, low-strength nitrogen, and temperature. Process Saf Environ Prot 194:1409-1420.
- Pan C, Wu J, Xu D, Guo L, Chen W, Wang J, Wu C, Zheng P, Zhang M (2025) Nitrogen removal function and its supporting properties of anammox granular sludge: A review. Chem Eng J 518:164742.
- Talan A, Tyagi RD, Drogui P (2021) Critical review on insight into the impacts of different inhibitors and performance inhibition of anammox process with control strategies. Environ Technol Innov 23:101553.
- Tang CJ, Duan CS, Yu C, Song YX, Chai LY, Xiao RY, Wei ZS, Min XB (2017) Removal of nitrogen from wastewaters by anaerobic ammonium oxidation (ANAMMOX) using granules in upflow reactors. Environ Chem Lett 15:311-328.
- Tao JS, Xing JL, Wang DP, Sheng BB, Meng FG (2020) Deciphering the genesis of anammox granular sludge floating from the perspective of microbial community. J Water Process Eng 36:101265.
- Waki M, Tokutomi T, Yokoyama H, Tanaka Y (2007) Nitrogen removal from animal waste treatment water by anammox enrichment. Bioresour Technol 98:2775-2780.
- Wang R, Liu JJ, Zhang Q, Li XY, Wang SY, Peng YZ (2024) Robustness of the anammox process at low temperatures and low dissolved oxygen for low C/N municipal wastewater treatment. Water Res 252:121209.
- Wang WG, Wang H, Jiang ZW, Wang YY (2020b) Visual evidence for anammox granules expanding their size by aggregation of anammox micro-granules. Sci Total Environ 745:141052.
- Wang WG, Yan Y, Zhao YH, Shi Q, Wang YY (2020a) Characterization of stratified EPS and their role in the initial adhesion of anammox consortia. Water Res 169:115223.
- Xiong YT, Li SY, Liu SY, Qiao LL, Chen YP, Yan P (2025) Enhancing the adaptation of anammox bacteria at low temperature: Alleviating the oxidative damage to anammoxosome membrane by ascorbic acid. Chem Eng J 505:159180.
- Xu RZ, Cao JS, Feng G, Luo JY, Feng Q, Ni BJ, Fang F (2022) Fast identification of fluorescent components in three-dimensional excitation-emission matrix fluorescence spectra via deep learning. Chem Eng J 430:132893.
- Yang T, Wang X, Chen H, Wang M, Wang Y, Chen H, Dai H (2024) Transcriptome analysis expands underlying mechanisms of quorum sensing mediating heterotrophic nitrification-aerobic denitrification process at low temperature. Bioresour Technol 414:131581.
- Zekker I, Raudkivi M, Artemchuk O, Rikmann E, Priks H, Jaagura M, Tenno T (2020) Mainstream-sidestream wastewater switching promotes anammox nitrogen removal rate in organic-rich, low-temperature streams. Environ Technol 1-23.
- Zhang J, Cao Y, Chen J, Zhang L, Wu W, Li J, Liu Y, Gai T, Yu J, Zhang H (2025) Important roles of immobilization in improving low-temperature nitrogen removal of cold-shock Anammox sludge and related mechanism. Environ Technol Innov 38:104170.
- Zhang JH, Miao YY, Zhang Q, Sun YW, Wu L, Peng YZ (2020) Mechanism of stable sewage nitrogen removal in a partial nitrification-anammox biofilm system at low temperatures: Microbial community and EPS analysis. Bioresour Technol 297:122459.
- Zhang M, Wang S, Bi Y, Meng F, Wang D, Qiu C, Wang C, Yu J (2022a) Enhanced nitrogen removal of single stage partial nitritation anammox system by glycine betaine addition at low temperature: Performance and mechanism. J Water Process Eng 49:102959.
- Zhang S, Lai LY, Wang TX, Jin WL, Yi RR, Chen DZ, Jin RC, Yang GF (2024) Response of anammox to organics with different degradation characteristics and exposure time: Performance, sludge characteristics and bacterial community. Sci Total Environ 951:175650.
- Zhang Y, Wu X, Cai J, Chen M, Zhang J, Shao S, Zhang Y, Ma Y, Wang Q (2023) Transposon insertion sequencing analysis unveils novel genes involved in luxR expression and quorum sensing regulation in Vibrio alginolyticus. Microbiol Res 267:127243.
- Zhang Y, Zhang J, Li J, Wei P, Luo R, Han H (2022b) Fast start-up of ANAMMOX biofilm processes at low temperatures by economical quorum sensing regulation: The importance of endogenous N-acyl-homoserine lactones from enhanced inoculated sludge. Environ Res 214:114097.
- Zhao Y, Liu S, Jiang B, Feng Y, Zhu T, Tao H, Tang X, Liu S (2018) Genome-centered metagenomics analysis reveals the symbiotic organisms possessing ability to cross-feed with anammox bacteria in anammox consortia. Environ Sci Technol 52:11285-11296.
- Zhu G, Wang S, Ma B, Wang X, Zhou J, Zhao S, Liu R (2018) Anammox granular sludge in low-ammonium sewage treatment: not bigger size driving better performance. Water Res 142:147-158.
Table 1 Operation conditions at each operation stage in different anammox reactors
|
Reactor |
Operational stages |
Operationdays (d) |
HRT(h) |
Influent (mg L-1) |
NLR (kg m-3 d-1) |
NRR (kg m-3 d-1) |
Average temperature (℃)* |
Temperature changing (℃)* |
||
|
NH4+-N |
NO2--N |
|||||||||
|
R1 |
I |
I-1 |
1-58 |
1.15 |
99.8±2.5 |
104.9±2.30 |
4.28±0.09 |
3.82±0.10 |
26.9±0.4 |
26-27 |
|
I-2 |
59-84 |
1.15 |
97.0±1.8 |
103.1±1.3 |
4.34±0.06 |
3.85±0.07 |
24.2±1.2 |
Gradually decreasing from 26 to 23 ℃ |
||
|
Ⅱ |
85-134 |
1.15 |
98.0±2.85 |
103.3±1.97 |
4.22±0.05 |
3.64±0.11 |
28.3±0.5 |
27-29 |
||
|
Ⅲ |
135-152 |
0.96 |
101.9±1.95 |
102.3±1.68 |
5.12±0.06 |
4.08±0.19 |
||||
|
Ⅳ |
153-182 |
0.96 |
112.1±13.77 |
111.4±12.75 |
5.63±0.70 |
4.83±0.61 |
||||
|
R2 |
Ⅱ |
85-134 |
1.15 |
98.0±3.00 |
104.8±2.91 |
4.25±0.06 |
3.26±0.34 |
20.6±2.5 |
Gradually decreasing from 23 to 15 ℃ |
|
|
Ⅲ |
135-152 |
0.96 |
104.3±1.98 |
100.0±1.03 |
5.13±0.05 |
3.16±0.38 |
15.0 |
15 ℃ |
||
|
Ⅳ |
153-182 |
0.96 |
74.4±2.78 |
71.2±1.73 |
3.65±0.08 |
2.55±0.27 |
16.1±2.5 |
Fluctuation between 11 and 20 ℃ |
||
*Water temperature in reactor.
Table 2 Granular size distribution of anammox granular sludge in both reactors
|
Reactor |
Characteristics |
>2 mm |
1-2 mm |
0.5-1 mm |
0.25-0.5 mm |
<0.25mm |
|
R1 |
SS (mg g-1) |
0.51 |
0.07 |
0.11 |
0.10 |
0.14 |
|
VSS (mg g-1) |
0.40 |
0.05 |
0.07 |
0.03 |
0.06 |
|
|
Proportion (%) |
54.65 |
7.79 |
12.21 |
10.33 |
15.02 |
|
|
VSS/SS (%) |
78.43 |
70.00 |
63.63 |
30.00 |
42.86 |
|
|
R2 |
SS (mg g-1) |
0.45 |
0.10 |
0.05 |
0.08 |
0.17 |
|
VSS (mg g-1) |
0.35 |
0.06 |
0.03 |
0.02 |
0.05 |
|
|
|
Proportion (%) |
52.49 |
11.64 |
6.34 |
9.68 |
19.85 |
|
|
VSS/SS (%) |
78.54 |
60.00 |
60.00 |
25.00 |
29.41 |
Competing interest reported. The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
{kind=link}