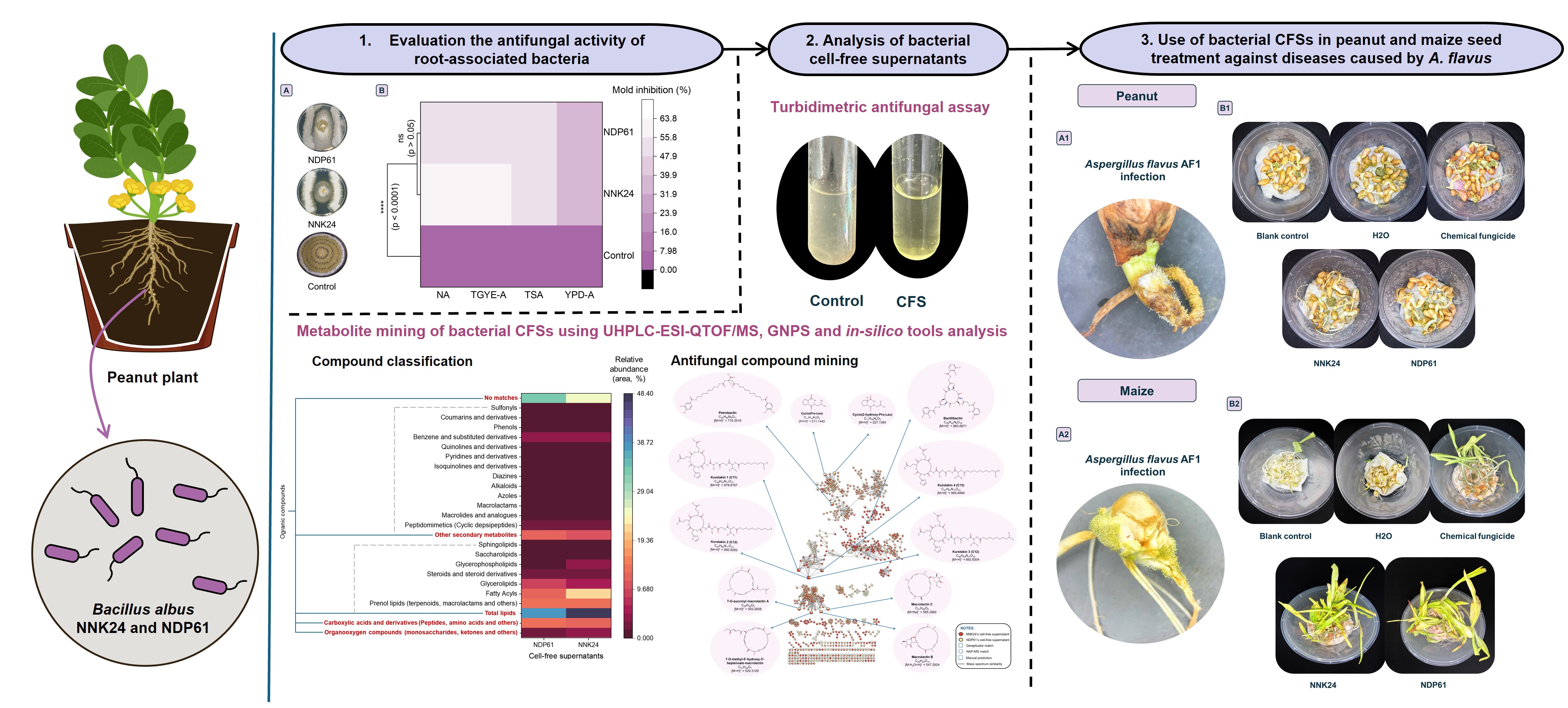
4.2. Antifungal metabolites in rhizobacterial CFSs
To outcompete their ecological competitors, bacteria typically employ diverse strategies, including the secretion of cell-wall degrading enzymes and/or secondary metabolites. Many antagonistic bacteria produce hydrolytic enzymes such as chitinase, β-glucanase, and protease, which play a crucial role in suppressing fungal growth [45]. However, in both examined CFSs, these essential fungal cell wall-degrading enzymes - namely chitinase, glucanase, and protease - were notably absent despite the addition of fungal biomass as enzyme inducers. Therefore, the antifungal activity of both CFSs was likely reliant on their secondary metabolites. Similarly, in the fermentation broth of Paenibacillus polymyxa HT16, the addition of chitin and β-glucan as inducers did not result in the presence of cell wall-degrading enzymes against fungal pathogens. However, antifungal low molecular weight compounds were detected in the culture broth supplemented with fungal biomass [46]. Another study indicated that bacteria release secondary metabolites upon interaction with fungal pathogens. For instance, rhizosphere B. subtilis was found to produce significant levels of small molecular weight compounds, effectively inhibiting the growth of competitors such as Setophoma terrestri [47]. Moreover, when exposed to phytopathogen biomass, Trichoderma asperellum produced various secondary antifungal compounds [48]. Co-cultivation of microbes is a well-known method to activate silent biosynthetic gene clusters for novel metabolite production, even when a member microbe is autoclaved [49].
The surfactant properties of bacterial cell-free supernatants (CFSs) suggest the presence of compounds with amphiphilic structures [50]. Bacillus spp. are known to synthesize many low molecular weight biosurfactants such as lipopeptides and glycolipids, where peptides or disaccharides are conjugated with long-chain fatty acids or hydroxy fatty acids [51]. These biosurfactants interact with cell membranes and can disrupt the membranes of various fungal species [52].
The composition of the CFSs, characterized by a high proportion of fatty acyls, glycerolipids, prenol lipids, cyclic depsipeptides, macrolactams, and macrolides, elucidated their surfactant nature. Several putative antifungal compounds were identified from CFS spectral data using both in-silico tools and manual predictions.
Firstly, the diketopiperazine cyclo(Pro-Leu), a compound produced by B. cereus, has demonstrated strong inhibition of mycelia growth in phytopathogenic A. flavus and A. niger, thereby impacting aflatoxin production [53].
Secondly, four macrolides from the macrolactin family were detected in both CFSs, including 7-O-succinyl macrolactin A, 7-O-methyl-5'-hydroxy-3'-heptenoate-macrolactin, macrolactin C, and macrolactin B. Remarkably, 7-O-succinyl macrolactin A, derived from B. amyloliquefaciens ELI149, exhibits robust antifungal activity against various significant filamentous fungal phytopathogens [54]. Additionally, macrolactin B and C, alongside 7-O-methyl-5'-hydroxy-3'-heptenoate-macrolactin, have been identified in B. subtilis MTCC10403 and Bacillus sp. AH159-1, respectively, showcasing broad-spectrum antibacterial activity [54, 55]. While their antifungal activity against plant phytopathogens remains unrecorded, a structurally similar compound, macrolactin R (also glycosylated similar to B and C), impacted the membrane structure of Botrytis cinerea, resulting in changes in membrane permeability, protein and nucleic acid leakage, and ultimately, cell death [56].
Thirdly, catecholate siderophores of Bacillus cereus group, petrobactin and bacillibactin, found in both CFSs, exhibit remarkable iron affinity, competitively inhibiting the growth of plant pathogens with less efficient iron-uptake systems [57, 58]. Bacillibactin produced by B. amyloliquefaciens MBI600 has been demonstrated to inhibit the growth of various fungi, including A. flavus, in iron-containing media. This suggests that its antibiotic function relies not only on iron scavenging to starve competing species but also on direct antibiosis against phytopathogenic fungi [59]. Additionally, microbial siderophores are recognized as a potential source of iron for plant survival and growth [60].
Finally, lipopeptides kurstakins (NRPs), identified in the CFSs of both NNK24 and NDP61 strains, have been previously reported to be linked to fatty acids ranging from C10 to C14, which may be β-hydroxylated or not, with two isoforms (n-, iso-). They are partially cyclic, forming a lactone bond between Ser/4 and the C-terminal Gln/7, and may also be linear [61, 62]. These amphiphilic lipopeptides act as biosurfactants, essential for swarming and biofilm formation, preventing the colonization and growth of competing microorganisms, and having a high affinity for cell membranes, through which they can form pores [20, 61, 63]. Consequently, they exhibit notable antimicrobial activities, including against various fungi. For examples, purified kurstakins 1–4 (isoC11, nC12, isoC12, isoC13), produced by B. thuringiensis kurstaki HD-1, demonstrating antifungal activity against Stachybotrys chartarum [64]. Subsequently, kurstakins (C10, C11, 12) were found to be produced by several biocontrol agents Bacillus spp. and Enterobacter cloacae C3 [62, 65]. Multiple kurstakin producers, such as B. cereus, B. thuringiensis, and B. pumilus, inhibited a certain number of fungi [66].
Taken together, it becomes evident that the majority of the identified compounds interacted with fungal cells. This observation leads us to hypothesize that the presence of fungal cells may prompt bacteria to develop strategies for producing various antifungal compounds that act through different mechanisms.
4.3. Potential to use rhizobacterial CFSs as bioprotection agents in agriculture
The infection of peanut and maize seeds with aflatoxigenic Aspergillus flavus AF1 during the germination experiment highlighted the severe impact of mycotoxigenic molds on seedling health. In both crops, the proliferation of mold mycelia and conidia, coupled with the formation of necrotic lesions on cotyledons, hypocotyls, and roots, inevitably leads to reduced productivity. The persistence of conidia throughout the plant life cycle can result in aflatoxin contamination in harvested products and during storage.
This study found that even in the blank control (seeds without A. flavus AF1 infection and without antifungal treatment), symptoms of A. flavus disease appeared on seedlings, indicating the presence of seed-borne A. flavus strains. This finding aligns with numerous reports on A. flavus in peanuts and maize [67, 68].
Treatment of seeds with CFSs of Bacillus albus NNK24 and B. albus NDP61 significantly mitigated the harmful effects caused not only by A. flavus AF1 but also by other seed-borne A. flavus strains that were indistinguishable from AF1 in peanut and maize seedlings, similar to the commercial fungicide Cruiser plus 312.5FS. This was evidenced by a notable reduction in Disease Incidence (DI) and Disease Severity Index (DSI). Moreover, these treatments promoted seedling growth, especially in peanuts, as indicated by various growth parameters, particularly the Vigor Index (VI).
However, the disease control efficacy of Cruiser Plus 312.5FS at the recommended concentration was limited (50.3–57.8%) in both seedlings. This limitation could be due to the potential resistance to the commercial fungicide despite containing two active ingredients, Difenoconazole and Fludioxonil, which have different antifungal mechanisms. This finding underscores the need for ongoing research into novel antifungal agents.
Using complex microbial metabolites, such as CFSs or their crude solvent extracts, to control phytopathogens offers several advantages. This approach avoids the safety concerns associated with using whole cells and harnesses the synergy of multiple antimicrobial compounds, thereby reducing the likelihood of early resistance development. For example, B. subtilis culture filtrate has shown inhibitory effects on A. flavus growth on peanuts [69]. Similarly, crude extracts from the CFSs of Bacillus sp. 16060, Serratia marcescens SE1, and S. marcescens SE2 have demonstrated antifungal activity against A. flavus in peanut seedlings [25]. In addition to controlling A. flavus, Bacillus extracts containing a complex mixture of antifungal compounds can reduce other plant fungal diseases [70]. As a recent trend, microbial CFSs are being explored as biostimulants and biocontrol agents in sustainable agricultures [71–74]. Furthermore, our previous study showcased the potent antifungal activity of bacterial cells of PGPR B. albus NNK24 and NDP61 in peanut seeds and plants [10]. Typically, bacterial endophytes confer various benefits upon their hosts [75]. Our study emphasizes the potential of utilizing endophyte CFSs to combat A. flavus not only in their host peanut but also in maize. This could significantly expand the application of bacterial products in sustainable agriculture, providing a safer and more effective alternative to chemical fungicides.
{kind=link}